Article Text

Abstract
Aim To describe the histological changes in the optic nerve (ON) after experimental segregation of cerebrospinal fluid (CSF).
Methods In seven sheep, a silicone band was placed around one ON to compress the subarachnoid space (SAS) surrounding the nerve, thus blocking the flow of CSF without compressing the ON itself. After 4 or 21 days, both the ligated and untouched ONs were removed and evaluated histologically.
Results All treated ONs showed marked loss of axons, destruction of myelin and swelling of meningoepithelial cells, most pronounced in the proximal ON adjacent to the globe at the location most distant to the ligature. There was no significant difference in histological findings between the ONs that were ligated for 4 days and those with 21 days of ligature.
Conclusion CSF segregation in the ON by blocking the SAS leads within 4 days to severe nerve damage. The increasing severity of these changes with increasing distance from the site of the ligature argues against simple pressure- or microperfusion-dependent effects and supports the hypothesis that interruption of CSF flow in the SAS of the ON can produce damage due to a change of CSF flow and content.
- Axons
- cerebrospinal fluid
- ligature
- myelin
- optic nerve
- papilloedema
- segregation
- optic nerve
Statistics from Altmetric.com
Introduction
Throughout its entire length, the optic nerve (ON) is surrounded by cerebrospinal fluid (CSF). Intracranially, the CSF simply surrounds and bathes the nerve within the basal cistern; however, within the orbit and optic canal, the nerve is covered by a sheath that consists not only of pia-arachnoid but also of dura, and the CSF thus is confined by the sheath. Studies measuring the concentration of lipocaline-like prostaglandin D synthase (L-PGDS), also known as β-trace protein, in the spinal CSF and the CSF in the subarachnoid space (SAS) of the ON demonstrate a concentration gradient in patients with normal-tension glaucoma and in patients with idiopathic intracranial hypertension (IIH).1 In addition, the influx of contrast-loaded CSF from the intracranial CSF into the SAS of the ON is markedly reduced in the same group of patients, thus supporting the concept of a segregation of ON CSF in such patients.2 Indeed, isolation of the SAS of the ON from the spinal and intracranial CSF spaces resulting in impaired CSF communication between the intracranial CSF and the CSF in the SAS surrounding the orbital and canalicular segments of the ON may be responsible for the occurrence of unilateral or highly asymmetric swelling of the optic discs in some patients with increased intracranial pressure.1–7
CSF is produced primarily by the choroid plexus epithelium, but also by meningoepithelial cells (MECs) that line the surface of the SAS and express L-PGDS.8 Biochemical studies of CSF sampled from the SAS of the ON during optic nerve sheath fenestration in patients with IIH, optic nerve sheath meningioma, posterior ischaemic optic neuropathy and normal-tension glaucoma demonstrate a concentration gradient of L-PGDS as well as other proteins between the SAS of the ON and the CSF sampled during lumbar puncture from the spinal SAS.1 2 7 These findings are consistent with other investigations of CSF composition in different parts of the SAS under certain pathological circumstances that also show concentration gradients of mainly brain-derived CSF proteins.9–11 The aim of this study was to create segregation of the CSF surrounding the ON from the intracranial CSF via a ligature, causing impairment of CSF communication in the SAS of the ON, in order to investigate the influence of CSF segregation on MECs, axons and myelin of the ON.
Materials and methods
Animals
Seven healthy female Dorper sheep (age 1–1.5 years, weight 30–35 kg) were selected. Animal facilities were maintained according to current guidelines for animal research provided by the United States National Institutes of Health (NIH) and the Association for Research in Vision and Ophthalmology (ARVO). Cefazolin 1000 mg was given by intravenous injection prior to surgery and followed by intramuscular injection of Procaine Penicillin G and dihydrostreptomycin sulphate once daily for 3 days. Carprofen was injected subcutaneously before the procedure at 4 mg/kg and for three following days, once every 24 h. The sheep were premedicated with intramuscular injections consisting of xylazine, 0.1 mg/kg, and ketamine, 10 mg/kg. Induction of anaesthesia was by intravenous injection of ketamine and midazolam, followed by intubation. Maintenance of anaesthesia was with isoflurane 1.5–2% combined with O2 100%. A nasogastric tube was inserted to prevent gas accumulation. Intravenous lactated Ringer's solution supplemented with bicarbonate was infused during surgery.
Surgical procedure
Following subcutaneous injection of bupivicaine at the incision site to achieve local anaesthesia, a skin incision was made over the temporo-zygomatic bone and carried down using sharp dissection and cautery until the temporo-zygomatic bone was reached. The bone was then removed using an oscillating bone saw to access the orbit. The lateral rectus muscle was myectomised to access the intraconal portion of the orbit. Blunt dissection was used to visualise the ON and carefully separate it from surrounding tissue. An elastic silicone band (Silikon Ptosis Probe, LLOuQuet+Guerin S3.1000; FCI Ophthalmics, Issy-les-Moulineaux, Cedex France) was placed around the portion of the nerve as close as possible to the optic canal, well posterior to the ciliary arteries. The silicone band was then tightened loosely so as to compress the SAS without actually compressing the nerve itself. After meticulous haemostasis was achieved, deep tissues were closed with absorbable sutures (Vicryl 3-0; Ethicon, Johnson & Johnson Medical, Langhorne, Pennsylvania, USA) and the skin incision was closed with Prolene sutures (Prolene 4-0; Ethicon, Johnson & Johnson Medical). A sterile dressing was placed over the incision site.
Four (n=3) or 21 (n=4) days after surgery, the sheep were anaesthetised using the protocol described above, the operated orbit was re-opened, and the globe and the entire orbital portion of the ON were removed leaving the silicone band in place. The removed tissue was placed in 3% paraformaldehyde and 2% sucrose in phosphate-buffered saline (PBS). In three sheep, the contralateral orbit was then opened using the same temporo-zygomatic approach as on the other side, and the globe and orbital portion of the ON were removed and processed in the same way as the other side, serving as controls. The sheep were then euthanised with isoflurane and intravenously injected sodium pentothal.
Histological procedures
Electron microscopy
After fixation with 3% paraformaldehyde (Nr. 76240; Sigma, Buchs SG, Switzerland) and 2% sucrose (Nr. 102745C; Chemie Brunschwieg, Basel, Switzerland) in PBS (Nr. BR0014G; LAB, Basel, Switzerland), tissue for electron microscopic examination was postfixed, first with 2% glutaraldehyde (Nr. 16210; VWR, Dietikon, Switzerland) and 0.8% paraformaldehyde in 50 mM sodium cacodylate buffer (pH 7.3) (Nr. 8.20670.0250; Merk, Darmstadt, Germany) and next, with 1% osmium tetroxide (OsO4) in 50 mM sodium cacodylate buffer (pH 7.3). The tissue then was dehydrated in an ethanol series (Nr. 02860; Fluka, Buchs SG, Switzerland) and embedded in epon (Nr. 2104502; Catalys, Duebendorf, Switzerland). Ultrathin sections of 50 nm were contrasted with uranyl acetate (Nr. 94260; Fluka) and lead citrate (Nr. 15334; Fluka) and studied with a CM-100 transmission electron microscope (Philips, Leiden, the Netherlands). For scanning electron microscopy, postfixed samples were dehydrated in an acetone series (Nr. 00570; Fluka), critical point dried, sputter-coated with 10 nm gold and imaged with a VP6360 scanning electron microscope (Jeol, Tokyo, Japan). Sections were analysed qualitatively with respect to organisation of myelin, axons and MECs in both the ligatured and control ONs.
Results
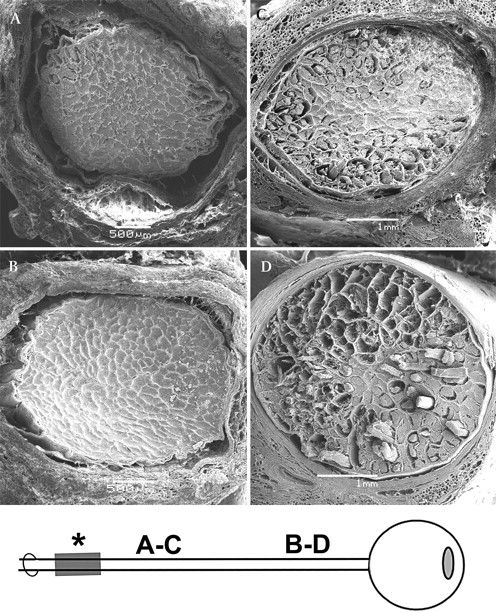
All ligatured ONs showed marked loss of axons by both light microscopy (figure 1) and scanning electron microscopy (figure 2) compared with control ONs. Axon loss was minimal adjacent to the site of the ligature (figure 2C), but became more pronounced with increasing distance distal to the site of the ligature, with maximal loss seen in the immediate retrobulbar portion (post-laminar portion) of the ON (figure 2D). Control ONs showed no evidence of axon loss at any point along the course of the ON (figures 2A,B).

Light microscopic sections of the control and ligated optic nerves (ON) (bulbar region, proximal, adjacent to globe). Trichrome Masson–Goldner stain, 20 μm sections. (A) Contralateral ON (serving as a control) showing normal axon bundles and normal width of the subarachnoid space (SAS). Note the normal width of the SAS and normal dimensions of meninges in untreated nerve. (B) Ligated ON displaying marked destruction of nerve fibre bundles. Note the narrowing of the SAS due to swelling in the arachnoidal/dural layer of the meninges. Note the swelling of arachnoidal meningeal structures with consequent narrowing of the SAS.
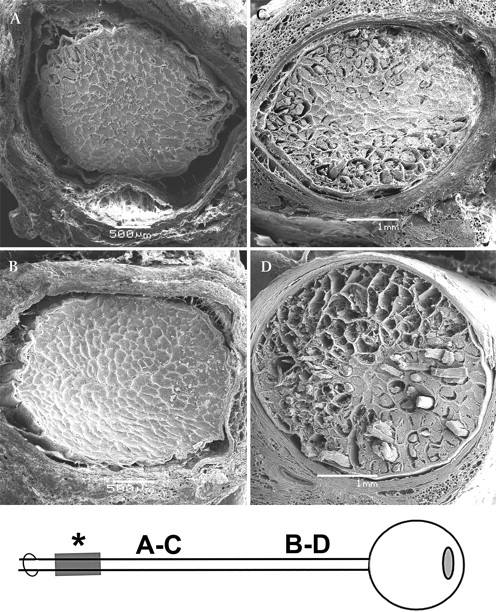
Scanning electron microscopy of cross-sections of normal (untreated side as control) (A, B) and ligatured (C, D) optic nerves (ON). (A, C) posterior (distal) portion; (B, D) anterior (proximal, adjacent to globe) portion. (A) Note the normally wide subarachnoid space (SAS) in the posterior portion of a control ON. (B) In anterior part of a control ON, note normally narrow SAS (this contrasts to human ON). (C) In posterior part of an ON that was ligatured for 4 days, note extensive myelin damage in the periphery of the nerve with few defects visible in its centre. (D) In anterior part of an ON that was ligatured for 4 days, note massive peripheral and central axon and myelin destruction.
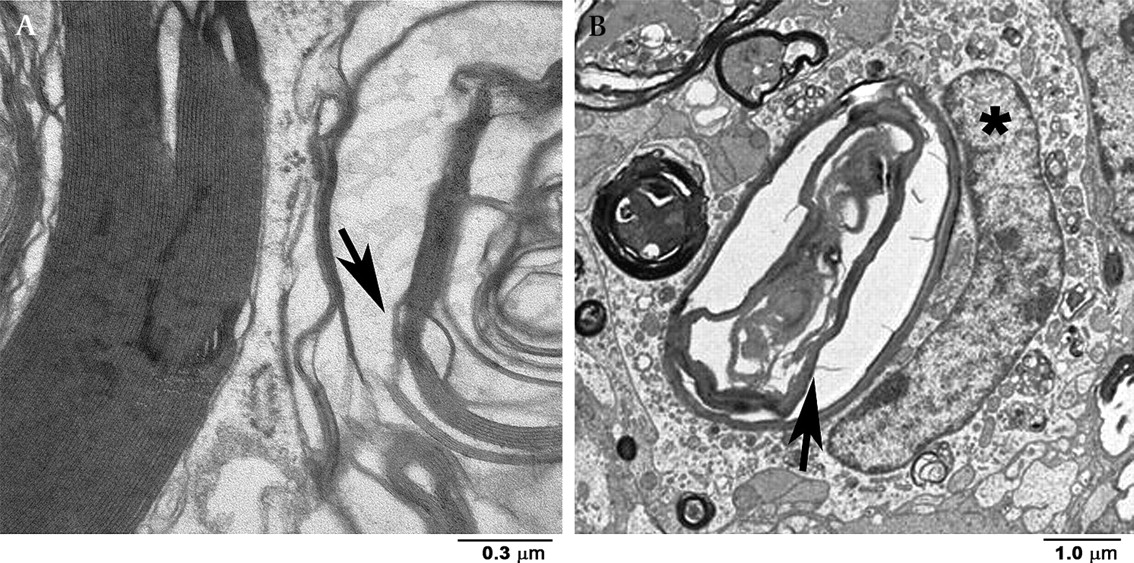
Transmission electron microscopy (figure 3) revealed substantial damage to myelin sheaths in the ligatured ONs, most prominent in the retrobulbar portion of the nerve adjacent to the globe (figure 3D). At some sites, intact myelin sheaths were adjacent to disarranged myelin sheaths (figures 3D and 4A). In contrast to the axons and their myelin sheaths, the cellular components of the ligatured ONs (oligodendrocytes and astrocytes) appeared relatively normal in both number and appearance by transmission electron microscopy (figure 4B). In all ligatured ONs, MECs surrounding the nerves were markedly swollen (figure 3C) compared with the MECs surrounding control ONs (figure 3A). The swelling of the MECs seemed to be associated with enlargement of intracellular structures (figure 3C). There was no significant difference in histological findings between the ONs that were ligatured for 4 days and those with 21 days of ligature.

Transmission electron microscopy of optic nerves (ON) without (A, B) and with (C, D) ligature-induced cerebrospinal fluid segregation. Bulbar region (retrobulbar portion, proximal, adjacent to globe). (A) Thin meningoepithelial cells (MECs) (arrow) line the surface of the meninges in the control ON. Asterisk, subarachnoid space. (B) Cross-section from a control (non-ligatured) ON shows normal-sized axons surrounded by intact myelin sheaths. (C) Cross-section from an ON ligatured for 21 days shows enlarged MECs (arrow). Note slight enlargement of intracellular structures (double arrow). (D) Cross-section from an ON ligatured for 21 days shows destroyed myelin sheaths and massive axonal loss.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Transmission electron microscopy in an optic nerve (ON) ligatured for 21 days. (A) Destroyed myelin sheath. Note adjacent intact myelin sheath on the left. (B) Nucleus of oligodendrocyte (asterisk) next to a destroyed axon (arrow) in a 21-day-ligatured ON.
Discussion
Until recently, CSF was believed to flow freely through all CSF spaces, associated with a homogeneous protein concentration and pressure.15 16 Recent work in humans has shown that this assumption is not always true, at least for patients with papilloedema and normal-tension glaucoma.1 2 A comparison between CSF from the spinal meninges and the CSF in the SAS of the ON in these patients demonstrates a significant concentration gradient of L-PGDS protein and a reduced influx of contrast-loaded CSF into the SAS of the ON from the basal cisterns.1 2 Both findings are consistent with a reduced circulation of CSF in the perioptic space.
To induce CSF segregation, we used an elastic silicone band around the ONs, being careful not to compress the ON itself but only the SAS. The impressive loss of axons and destruction of myelin, as well as the marked swelling of MECs of the arachnoid as well as the trabeculae and septae in the SAS, suggests that impaired CSF circulation can lead to ON damage. On cross-sections of the ligated ON, we found an uneven pattern of axon loss with maximum damage in the portion of the nerve adjacent to the globe; that is, at the site most remote from the ligature. The least affected area was at the site of the ligature, where damage was confined to the periphery of the nerve in a circular pattern, whereas the damage to the nerve distal to the ligature involved nearly the entire cross-section. Thus, although the ligature may have caused minor damage by direct compression or surgical trauma, the major damage remote from the ligature was caused by another process that we believe to be related to CSF segregation.
The technique we applied in this study was somewhat similar to that previously used by Wirtschafter et al, who placed a silk suture around the ON just posterior to the globe to investigate the pathophysiology of papilloedema without increasing intracranial pressure.17 These investigators found evidence of damage to ON axons characterised by extensive but highly variable axon diameters, a finding similar to that of our study, in which we also found an uneven distribution of axon loss in different locations in the ON. However, our model differs from that of Wirtschafter et al in that they used non-human primates, whereas we performed our experiments in sheep, and their ligature was a silk suture placed beneath the posterior ciliary vessels surrounding the ON adjacent to the globe, whereas our ligature was an elastic silicone band placed posteriorly around the nerve near the optic canal, well behind the ciliary vessels. In addition, we applied only a little tension to the silicone band with the intention of compressing only the SAS and not the ON itself. As primates may differ in their immune responses from other mammals, it is possible that the silk suture used by Wirtschafter et al produced a more intense and rapid inflammation in the ON than our ligature. This could explain the extensive changes (eg, axon swelling and an increased number of mitochondria in the optic disc) they observed within just 2 days after placement of the suture.17 We believe that by using an elastic silicone band placed loosely around the more posterior aspect of the orbital portion of the ON, we created a less destructive and more controlled environment, leading to interruption of CSF communication in the SAS of the ON but not to stasis of axonal transport, as was the goal of the study by Wirtschafter et al. This difference in technique is also reflected in the fact that in our study, there were almost no fixation artefacts observed in the electron microscopy images in the studied portions of the ON, whereas in the study by Wirtschafter et al, numerous artefacts were present.17
We found no significant difference in light-microscopic or electron-microscopic findings between the nerves that were ligatured for 4 days and those in which the ligature remained for 21 days. This finding is consistent with the marked damage within 2 days described by Wirtschafter and colleagues17 and indicates that the major damage to the ON in our model occurred shortly after placement of the band and was not a progressive process, such as that which would be expected if the band were causing compression of the nerve.
There are at least four possible mechanisms that could explain the axonal damage in our model: (1) pure mechanical pressure-induced injury to axons; (2) vascular damage to the pial plexus surrounding the ON; (3) hydrostatic or hydrodynamic pressure; and (4) disturbed biochemical and metabolic function of the nerve from CSF segregation.
The strongest argument against a pure mechanical pressure-induced injury in our model is that in all of our specimens, the cross-sections most remote from the site of the ligature showed the most extensive damage. The ligature was placed as far from the globe as possible, but the cross-section in the immediate retrobulbar portion showed the most severe axonal damage, whereas the cross-sections in the area of the ligature showed only minimal damage. A local compressive mechanism causing the damage as shown in our model thus seems highly unlikely.
As far as vascular damage to the pial plexus surrounding the ON is concerned, the intraorbital ON in sheep is similar to that in humans, except that it lacks a central retinal artery.18 It is supplied by arterioles and capillaries that follow a centripetal pathway from a pial plexus that surrounds the ON. The central retinal artery and the ciliary arteries contribute little to the vascular supply of the nerve, as evidenced by the fact that neither central retinal nor ciliary artery occlusions occur in humans with posterior ischaemic optic neuropathy or optic nerve sheath meningioma. If the pial plexus were compressed by the ligature, we would expect the centre of the ON, being the most remote area from the feeding vessels, to be damaged first, a pattern of damage that is seen in some microvascular oculomotor nerve palsies that occur in patients with diabetes mellitus19 20 In our model, the opposite is the case; that is, the area affected at the site of the ligature is the periphery of the ON, not its centre, and this pattern of damage does not change over time.
The axonal damage seen in our model could be caused by hydrostatic/hydrodynamic pressure. Pressure measurements within the CSF surrounding the ON have been performed in human cadavers.21 We did not perform such measurements in our animals, because the SAS in sheep is rather narrow without a distended bulbar portion, and it is our belief that the relationship of the needle size that would be required to perform the measurements and the diameter of the SAS surrounding the ON would make it difficult to obtain reliable pressure readings. Furthermore, the SAS is not an open space but is divided by trabeculae and septae into separate small chambers containing CSF.22 A needle entering one of these chambers would thus provide information only about the pressure in that particular pocket and not necessarily throughout the SAS of the ON. Indeed, several attempts to create a model of CSF flow have been made by bioengineers, but none of these models can describe adequately CSF flow in a complex multi-chambered space such as the SAS of the ON.23 24 In addition, as the amount of CSF in the SAS is minute, the hydrostatic pressure is probably insignificant and would be so even if the volume of CSF in the SAS of the ON tripled. Having said this, without having performed pressure measurements in our study, we cannot exclude the possibility of fluid-derived pressure-induced damage on axons and myelin in our model.
We believe that disturbed biochemical and metabolic function of CSF due to segregation is the most likely, or at least the most important, mechanism for the ON damage seen in our model. Obstruction of CSF circulation in the SAS (ie, CSF segregation) could lead to a change in the biochemical composition of the CSF and to accumulation of certain biochemical active substances that could damage the ON. For example, it has been shown that depending on its concentration, L-PGDS, a protein present in the CSF, can interfere with ON metabolism in vitro through its effects on astrocytes and their mitochondria,25–27 and that fibroblast growth factor 2, also present in CSF, inhibits oligodendrocyte function and inhibits remyelination in vivo in monkeys.28 29 These findings indicate that CSF composition has an important impact on cellular functions within the central nervous system, as occurs in certain degenerative neurological conditions, such as normal-pressure hydrocephalus and Alzheimer's disease.30 31
In addition, a study in six patients with anterior ischaemic optic neuropathy, optic nerve sheath meningioma, idiopathic optic disc swelling (presumed from a local vascular cause), pachymeningitis, or idiopathic intracranial hypertension and highly asymmetric optic disc swelling following a cerebral haemorrhage, our group found an impressive concentration gradient (ratio L-PGDS concentration in the CSF taken from the SAS of the ON during optic nerve sheath fenestration divided by L-PGDS concentration in lumbar CSF) of L-PGDS between the spinal CSF and the CSF in the SAS of the ON of 450% (mean values) up to 685% (maximum values).1 This gradient is far greater than the gradient between the lumbar CSF and the intracranial CSF for brain-derived proteins described by Reiber.11 Taken together these data provide circumstantial evidence for CSF segregation in at least some disorders in which there is damage to the ON.
Finally, the finding that the most damaged area of the ON in our model was the immediate retrobulbar portion adjacent to the globe could be explained by the fact that this is by far the most metabolically vulnerable portion of the nerve. After passing through the lamina cribrosa, unmyelinated ON axons continue to lack myelination for a short distance. This unmyelinated, extraocular part of the ON contains the highest concentration of mitochondria in the nerve, and these mitochondria appear to play a crucial role in nerve function.32 33 They are transported from the retinal ganglion cells along axon-cytoskeletal structures to reach their destination.34 This distribution of mitochondria is thought to allow sufficient bioenergetic potential for undisturbed signal transmission in unmyelinated axons.32 33 If CSF accumulates in this vulnerable region of the ON, certain substances within the CSF such as L-PGDS could lead to loss of axons and destruction of myelin. Unfortunately, for the reasons stated above, we were not able to collect CSF for reliable chemical analysis in this study.
The mechanism by which CSF accumulates around the ON and produces a CSF segregation characterised by a change in concentration of certain, potentially harmful substances (eg, L-PGDS) is unclear but may involve the lymphatic vessels in the ON meninges that have been identified by both light and electron microscopy and have been shown to participate in CSF outflow pathways.35–42 As noted above, the SAS of the ON contains numerous septae and trabeculae that divide it into many small chambers.22 Although these chambers seem to communicate with each other in healthy subjects, disruption of this free ‘interchamber’ communication due to a local compressive or inflammatory process involving the meninges and especially the MECs lining the inner surface of the meninges, could lead to separation of these SAS chambers. Disruption of the CSF outflow via a lymphatic pathway in the meninges thus is one possible mechanism that could lead to such inflammation and subsequent CSF segregation. Further studies of CSF segregation optic neuropathy therefore should focus on CSF analysis, histological changes of lymphatic vessels in the ON sheath and changes of the optic disc (eg, papilloedema).
A potential weakness of this study is that we did not perform sham surgery on any of our animals, so we cannot say for certain if the surgery itself could have caused some or all of the changes seen in the ligated ONs at days 4 and 21. We consider this unlikely in view of both the location and the extent of the changes seen in those nerves. In addition, as identical surgery was performed in the sheep whose ONs were used as controls, we can at least state that there were no acute changes caused by the surgery in those nerves. It may be appropriate to perform sham surgery in subsequent studies.
Acknowledgments
The authors would like to thank the Department of Anatomy, University of Zurich, Switzerland, for the histological pictures (Therese Bruggmann and Gery Barmettler for excellent technical help in electron microscopy); the Animal Research Center, Hadassah University Hospital, Jerusalem; Israel for excellent animal-keeping facilities; and the Helmut Horton Foundation, Agno, Switzerland and the Guggenheim-Schnurr Foundation, Basel, Switzerland, for financial support of this basic science study without any formal legal requirements. Special thanks to P Groscurth for assisting with scanning electron microscopy.
References
Footnotes
Funding This study was supported by two private foundations: the Helmut Horton Foundation (Agno, Switzerland) and the Guggenheim-Schnurr Foundation (Basel, Switzerland).
Competing interests None.
Ethics approval This study was performed according to the Association for Research in Vision and Ophthalmology (ARVO) guidelines for animal research and was approved by the local ethical committee.
Provenance and peer review Not commissioned; externally peer reviewed.