Article Text

Abstract
Background and aim: Stevens–Johnson syndrome (SJS) and toxic epidermal necrolysis (TEN) are acute-onset mucocutaneous diseases induced by infectious agents and/or inciting drugs. Given the association between the onset of SJS/TEN and infections, the possibility that there is an association between SJS/TEN and a disordered innate immune response was considered. The first line of defence against infection is comprised of evolutionarily conserved sets of molecules, the Toll–like receptors (TLRs). TLR3 recognises double-stranded RNA associated with viral infections.
Methods: The Japanese single-nucleotide-polymorphism (JSNP) database reports 7 polymorphisms consisting of 7 SNPs in the human TLR3 gene; 3 of the 7 SNPs are coded in exon regions, (ie, 293248A/G, 293391A/G and 299698T/G), and the other 4 are coded in intron regions, (ie, 294440G/C, 294732C/T, 208036T/C and 298054C/T). These 7 SNPs were analysed in 57 Japanese patients with SJS/TEN with ocular surface complications and in 160 Japanese healthy controls.
Results: SNP 299698T/G and the genotype patterns of 293248A/A and 299698T/T were strongly associated with SJS/TEN.
Conclusion: The results suggest that polymorphisms in the TLR3 gene could be associated with SJS/TEN in the Japanese population.
- ds RNAs, double stranded RNAs
- RV, rhinovirus
- SJS, Stevens–Johnson syndrome
- SNP, single-nucleotide polymorphism
- TEN, toxic epidermal necrolysis
- TLR3, Toll-like receptor 3
Statistics from Altmetric.com
- ds RNAs, double stranded RNAs
- RV, rhinovirus
- SJS, Stevens–Johnson syndrome
- SNP, single-nucleotide polymorphism
- TEN, toxic epidermal necrolysis
- TLR3, Toll-like receptor 3
Stevens–Johnson syndrome (SJS) and the related disease toxic epidermal necrolysis (TEN) are acute multisystem inflammatory disorders of the skin and mucous membranes, including the ocular surface. They are commonly associated with infectious agents and/or an inciting drug.1 The annual incidence of SJS and TEN has been estimated as 0.4–1 and 1–6 cases per million persons, respectively,1,2 and the mortality is 3% and 27%, respectively.3 Although SJS and TEN are rare, they carry high morbidity and mortality, and often result in severe handicaps such as loss of vision. The rarity of cutaneous, mucosal and ocular surface reactions to drug therapies led us to suspect individual susceptibility. Therefore, we examined the possibility of a genetic predisposition towards SJS/TEN.
SJS/TEN is one of the most devastating ocular surface diseases, leading to corneal damage and loss of vision. The reported incidence of ocular complications in SJS/TEN is 50–68%.1,3 Ophthalmologically, in the acute stage, patients with SJS/TEN manifest vesiculobullous skin lesions, severe conjunctivitis and persistent corneal epithelial defects because of ocular surface inflammation. In the chronic stage, ocular surface complications such as conjunctival invasion into the cornea due to corneal epithelial stem-cell deficiency, symblepharon and ankyloblepharon, and in some instances keratinisation of the ocular surface, persist (fig 1A) despite healing of the skin lesions.4 We observed that >95% of the patients with SJS/TEN with ocular surface complications had lost their finger nails in the acute or subacute stage, and that some continue to have transformed nails even after healing of the skin lesions (fig 1B). In the current study, we focused exclusively on patients with SJS/TEN accompanied by ocular surface complications.
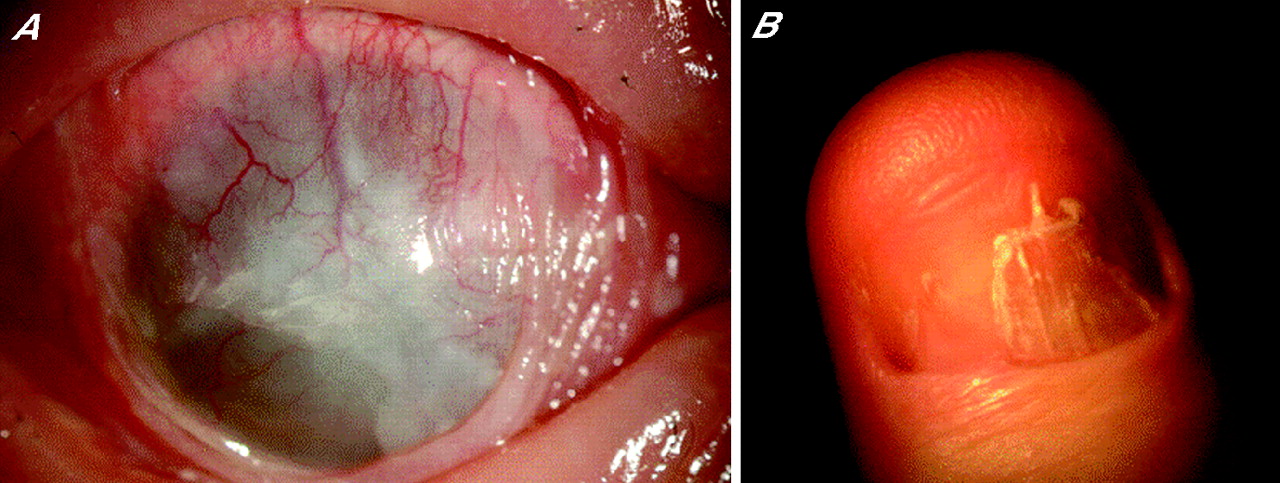
Patients with Stevens–Johnson syndrome/toxic epidermal necrolysis (SJS/TEN) with ocular complications. (A) Ocular surface complications such as conjunctival invasion into the cornea, symblepharon and ankyloblepharon, and sometimes keratinisation of the ocular surface, persist in some patients with SJS/TEN in the chronic stage. Conjunctival invasion results in severe vision loss. (B) Transformed fingernails of patients with SJS/TEN with ocular complications. Many patients with SJS/TEN with ocular complications lost their fingernails during the acute stage, and some continue to have transformed nails even after healing of the skin lesions. The photograph shows the thumbnail of a 26-year-old man 6 years after onset of the disease (chronic stage).
Drugs are probably the most widely accepted aetiological factor in SJS/TEN.5 In addition, patients with SJS/TEN often had prodromata, including non-specific fever, coryza and sore throat, that closely resemble upper respiratory tract infections commonly treated with antibiotics. These prodromata were evident from the clinical records of our patients with SJS/TEN. Mycoplasma pneumoniae was responsible for 5 of 17 cases of childhood SJS,6 and a viral aetiology involving herpes simplex, Epstein–Barr, cytomegalo-, and varicella zoster virus has been reported.7,8
Given the association between the onset of SJS/TEN and infections, and the opportunistic infection of ocular surfaces by bacteria such as methicillin-resistant Staphylococcus aureus or methicillin-resistant Staphylococcus epidermidis,9 we considered the possibility of an association between SJS/TEN and a disordered innate immune response. We postulated that viral infection and/or drugs might trigger a disorder in the host’s innate immune response, and that this event is followed by aggravated inflammation of the mucous membranes, ocular surface and skin.
The first line of defence against infection is comprised of evolutionarily conserved sets of molecules, the Toll-like receptors (TLRs). The triggering of TLRs results in the secretion of antibacterial peptides and proinflammatory cytokines. The inflammatory response results in the recruitment of cells of adaptive immunity to initiate clearance of the pathogens. TLR3 recognises double-stranded (ds)RNA associated with viral infections, a component of the life cycle of most viruses.10 As functional deterioration of TLR3 can predispose individuals to increased susceptibility to viral infections, the detection of TLR3 polymorphisms could yield critical information for risk assessment regarding susceptibility to microbial infections in the context of SJS/TEN.
To date, no reports are available on the genetic loci of TLR3 in subjects with SJS/TEN. Therefore, we performed single-nucleotide polymorphism (SNP) association analysis of the TLR3 gene, which maps to chromosome 4q35. The Japanese single-nucleotide polymorphism (JSNP) database reports seven polymorphisms consisting of seven SNPs in the human TLR3 gene: three of the seven SNPs are coded in exon regions ie, 293248A/G (rs.3775290, exon 4, silent SNP), 293391A/G (rs.3775291, exon 4, change SNP) and 299698T/G (rs.3775296, exon 2, UTR SNP), and the other four are coded in intron regions (ie, 294440G/C (rs.3775292, intron 3), 294732C/T (rs.3775293, intron 3), 208036T/C (rs.3775294, intron 2) and 298054C/T (rs.3775295, intron 2; fig 2).

{kind=link}
{kind=link}
Genomic organisation of the Toll-like receptor 3 (TLR3) gene on chromosome 4q35. The genomic organisation of the gene was derived from the Japanese single-nucleotide polymorphism (JSNP) database. Seven polymorphisms consisting of seven SNPs have been reported in the human TLR3 gene in the JSNP database; three of the seven SNPs are coded in exon regions (ie, 293248A/G (rs.3775290, exon 4, silent SNP), 293391A/G (rs.3775291, exon 4, change SNP) and 299698T/G (rs.3775296, exon 2, UTR SNP)), and the other four are coded in intron regions (ie, 294440G/C (rs.3775292, intron 3), 294732C/T (rs.3775293, intron 3), 208036T/C (rs.3775294, intron 2) and 298054C/T (rs.3775295, intron 2)).
We analysed these seven SNPs in 57 Japanese patients with SJS/TEN with ocular surface complications and in 160 healthy Japanese controls. We found that SNP 299698T/G and the genotype patterns of 293248A/A and 299698T/T are strongly associated with SJS/TEN.
MATERIALS AND METHODS
Patients
This study was approved by the institutional review board of the Kyoto Prefectural University of Medicine, Kyoto, Japan. All experimental procedures were conducted in accordance with the principles set forth in the Declaration of Helsinki. The purpose of the research and the experimental protocols were explained to all participants, and their prior written informed consent was obtained.
For SNP analysis, we enrolled 57 patients with SJS/TEN in the chronic or subacute phase; all presented with ocular surface complications. The diagnosis of SJS/TEN was based on a confirmed history of the acute onset of high fever, serious mucocutaneous illness with skin eruptions and involvement of at least two mucosal sites, including the ocular surface. The controls were 160 healthy volunteers. All participants and volunteers were Japanese, residing in Japan. The mean (SD) age of the patients involved in this study was 45.2 (17.5) years, and that of the controls was 36.2 (11.5) years. The ratios of men/women in the patient and control groups were 24/33 and 57/103, respectively.
SNP analysis
TLR3 SNP analysis was performed by sequencing from both sides, forward and reverse, to confirm the results carefully. For SNPs of TLR3, the PCR and sequence primers were 5′-TGGCTAAAATGTTTGGAGCA-3′ (sense) and 5′-GAAGAGGCTGGAATGGTGAA -3′ (antisense) for rs.3775290 and rs.3775291, 5′-CAGTTCTTTACTCCATCTCCGC-3′ (sense) and 5′-CCAAGGCTCTGGTAAGGGTG-3′ (antisense) for rs.3775292 and rs.3775293, 5′-TCACATGGCTTATCAAACACACAG-3′ (sense) and 5′-CATTGCTCTTCCTCAGATGCC-3′ (antisense) for rs.3775294 and rs.3775295, and 5′-TTACCTTCTGCTTGACAAAGGG-3′ (sense) and 5′-TGCATTTGAAAGCCATCTGC-3′ (antisense) for rs.3775296. All primers except for rs.3775290 and rs.3775291, were those recommended in the JSNP database. Genomic DNA was isolated from human peripheral blood at SRL (Tokyo, Japan). PCR amplification was done with DNA polymerase (Takara; Shiga, Japan) for 35 cycles at 94°C for 1 min, and annealing at 60°C for 1 min and at 72°C for 1 min on a commercial PCR machine (GeneAmp; Perkin-Elmer Applied Biosystems, Foster City, California, USA). The PCR products were reacted with BigDye Terminator V.3.1 (Applied Biosystems, Foster City, California, USA), and sequence reactions were resolved on an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems).
Statistical methods
Alleles as well as genotype patterns were counted manually. For Hardy–Weinberg equilibrium and statistical analysis to compare allelic and genotypic distributions, we used the χ2 test. The odds ratio (OR) with 95% CI was calculated using Labo Server software (World Fusion, Tokyo, Japan). Each allele and genotype pattern was assessed as an independent variable, and separate p values were calculated for each polymorphism. A p value of <0.05 was regarded as significant. In addition, the p values were corrected for the number of alleles tested (Bonferroni method).
RESULTS AND DISCUSSION
Table 1 shows a summary of our case–control association study on the seven SNPs genotyped to TLR3.
Genotype frequencies of toll-like receptor 3 single-nucleotide polymorphisms among Japanese patients with Stevens–Johnson syndrome/ toxic epidermal necrolysis and healthy controls
All but SNP 294732C/T were in Hardy–Weinberg equilibrium in both patients with SJS/TEN and controls (p>0.05). SNP 299698T/G showed a significant association under a recessive model (299698 T/G + G/G vs T/T, raw p value = 0.001, corrected p value = 0.007, OR = 0.22). However, when we corrected the p value for the number of alleles tested (n = 7), the results were no longer significant; SNP 293248A/G also showed a significant association under a recessive model (293248 A/G + G/G vs A/A, raw p value = 0.046, corrected p value = 0.322, OR = 0.46).
We also analysed the genotype patterns of SNPs 299698T/G and 293248A/G and found that these (293248A/A–299698T/T) were also strongly associated with SJS/TEN in Japanese patients (χ2 test, p = 0.001, OR = 5.5, 95% CI = 1.9 to 15.8) (table 2). This association was stronger than that observed for the single locus (299698T/G).
Pattern structures and frequency of single-nucleotide polymorphisms 293248A/G and 299698T/G
Our results suggest that polymorphisms in the TLR3 gene could be associated with SJS/TEN in the Japanese population. We hypothesised that viral infection and/or drugs might trigger a disorder in the host innate immune response, and that this event is followed by aggravated inflammation of the mucous membranes, ocular surface and skin. Genetic and environmental factors could play a role in an integrated aetiology of SJS/TEN.
Because the 299698T/G SNP, which showed a significant association with SJS/TEN, is encoded in the exon region, we consider it important to extend this study by performing expression and function analysis of the TLR3 protein with this SNP. According to the International HapMap project, the 299698T/G (rs.3775296) SNP exists not only in Japanese (G/G 0.386, G/T 0.500, T/T 0.114) but also in Han Chinese (G/G 0.659, G/T 0.295, T/T 0.046) and Caucasian (G/G 0.719, G/T 0.263, T/T 0.018) populations, indicating that it is important to examine TLR3 SNPs in non-Japanese populations.
TLR3 is involved in responses to dsRNAs.10 As rhinoviruses are a major cause of the common cold and the acute exacerbation of chronic obstructive pulmonary disease, the functional requirement for TLR3 in the host response against infection with live viruses, especially rhinovirus infection, has been proposed.11
The association documented here complements previous findings compatible with an unregulated innate immune response as an important pathophysiological condition in inflammatory ocular surface diseases.12–14 SJS/TEN could be the consequence of exposure of genetically susceptible individuals to specific environmental precipitants. A report from the US showed that the HLA (human leucocyte antigen)-B12 HLA-Bw44 antigen was significantly increased in Caucasian patients with SJS with ocular involvement.15 Analyses of patients with TEN in France also disclosed an association with HLA-B12 (HLA-Bw44).16 In Han Chinese, there was a very strong association between carbamazepine-induced SJS and the HLA-B*1502 allele.17 Elsewhere we reported that, in the Japanese, HLA-A*0206 was strongly associated with SJS/TEN, with ocular surface complications.18 These findings suggest that SJS/TEN is associated with a complex genetic-inheritance background, and that specific combinations of genes are required for disease-onset.
The pathophysiological mechanisms underlying the onset of SJS/TEN have not been fully established, although the involvement of immune mechanisms and altered drug metabolism has been suggested.19–23 Heretofore, it was not recognised that innate immunity plays a critical role in the pathophysiology of SJS/TEN, by way of bridging between the acute response to invading non-self molecules and chronic local immune inflammation.
We previously reported that while human corneal epithelium harbours messages for most TLRs, TLR3 is most highly expressed.12 In conjunctival, as in corneal, epithelium, TLR3 is the TLR with the highest expression level at the messenger RNA level (data not shown). We reported that the cell-surface TLR3 of human corneal epithelial cells responds to virus dsRNA-mimic polyI:C to generate proinflammatory cytokines and interferon β, and that the innate immune responses in human corneal epithelial cells differ from those in immune-competent cells.12 In the current study, we clarified the association between Japanese patients with SJS/TEN and TLR3 gene polymorphisms. This raises the possibility that abnormalities of TLR3 on the ocular surface might contribute to ocular surface inflammation such as SJS/TEN. In addition, the association between the onset of SJS/TEN and viral infections raises the possibility of an association between SJS/TEN and a disordered innate immune response.
The association between TLR polymorphisms and human diseases has been suggested. Polymorphisms in the TLR3 gene could be associated with type 1 diabetes in black people from South Africa.24 In the children of European farmers, there was a strong association between TLR2 polymorphisms and allergic diseases.25 Torok et al26 reported an association between a functional polymorphism in TLR4 and ulcerative colitis. The specific link between exposure to environmental triggers and the induction of a highly restricted autoimmune process remains to be detected, and the innate immune system could constitute a link between the environment and the adaptive immune system.
Acknowledgments
We thank Ms C Mochida for her technical assistance.
This study was supported in part by grants-in-aid for scientific research from the Japanese Ministry of Health, Labour and Welfare, the Japanese Ministry of Education, Culture, Sports, Science and Technology, CREST from JST, a research grant from the Kyoto Foundation for the Promotion of Medical Science, and the Intramural Research Fund of the Kyoto Prefectural University of Medicine.
REFERENCES
Footnotes
-
Published Online First 21 February 2007
-
Competing interests: None declared.