Article Text

Abstract
In this review, we aim to summarise key articles that explore relationships between the gut and ocular surface microbiomes (OSMs) and immune-mediated dry eye. The gut microbiome has been linked to the immune system by way of stimulating or mitigating a proinflammatory or anti-inflammatory lymphocyte response, which may play a role in the severity of autoimmune diseases. Although the ‘normal’ gut microbiome varies among individuals and demographics, certain autoimmune diseases have been associated with characteristic gut microbiome changes. Less information is available on relationships between the OSM and dry eye. However, microbiome manipulation in multiple compartments has emerged as a therapeutic strategy, via diet, prebiotics and probiotics and faecal microbial transplant, in individuals with various autoimmune diseases, including immune-mediated dry eye.
- Microbiology
- Ocular surface
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
The gut microbiome is composed of a plethora of bacteria that play important physiological roles in maintaining host health. First, certain microbiome constituents break down food molecules that are otherwise indigestible, such as plant carbohydrates that can be digested by the phylum Bacteroidetes.1 Second, the microbiome is involved in vitamin synthesis, such as folate production by the genus Bifidobacterium and vitamin K production by several species.2 Third, the gut microbiome helps regulate immune responses, balancing protection against pathogenic organisms while modulating inflammation intraintestinally and extraintestinally. For example, plasma cells in mucosal tissues, including the gut, produce large amounts of immunoglobulin A (IgA) that neutralise pathogenic bacteria and promote survival of commensal bacteria.3 In one study, germ-free mice (mice without a gut microbiome) harboured fewer IgA-producing plasma cells compared with control mice. However, plasma cell levels increased significantly after a microbiome was transplanted via faecal transfer from healthy wild-type mice, demonstrating the importance of the microbiome in regulating antibody production and immune responses.4 Furthermore, a healthy gut microbiome protects against infection by pathogenic bacteria, as evidenced by susceptibility to Salmonella typhimurium and Clostridium difficile in Swiss-Webster mice with reduced intestinal diversity.5 These are just a few examples of the important roles played by the gut microbiome in maintaining health, metabolic function and immunological responses.
Because the gut microbiome influences immune function, many studies have examined whether gut microbiome composition varies in individuals with autoimmune diseases, such as ankylosing spondylitis, Behcet’s and rheumatoid arthritis (RA) as compared with controls.6–8 In this review, we focus on relationships between gut microbiome composition and immune-mediated dry eye, a disease that often coexists with primary Sjögren’s, as well as secondary Sjögren’s (such as in the setting of RA) and graft-vs-host disease (GVHD). First, we describe the ‘normal’ gut microbiome and discuss connections between the gut microbiome and immune system. Next, we summarise data regarding gut microbiome compositional differences in individuals with immune-mediated dry eye and related diseases as compared with controls. Then, we briefly comment on interactions between the ocular surface microbiome (OSM) and dry eye, given that this is an area less well studied. Finally, we discuss strategies that can be used to alter the gut and OSMs and examine their potential as treatments for dry eye.
Methods
In this review, we summarise articles that discuss relationships between microbiomes and immune-mediated dry eye in Sjögren’s, as well as related diseases. Articles for this review were collected from the PubMed database using a non-systematic literature search. A mix of keywords was used to identify articles for this review including: ‘microbiome’, ‘dysbiosis’, and ‘commensal bacteria’ combined with terms such as ‘Sjögren’s’, ‘graft vs host disease’, ‘rheumatoid arthritis’, ‘autoimmune’, ‘dry eye’, ‘conjunctivitis’, ‘keratoconjunctivitis sicca’, ‘therapy’, ‘fecal microbial transplant’, ‘probiotics’, ‘prebiotics’ and ‘diet’. All published scientific articles were considered including original research, meta-analyses and systematic reviews. When multiple studies were available on the same topic, we chose articles with the most robust methodology that were pertinent to our review. All searches were limited to the English language. Overall, the search yielded 2325 articles on PubMed of which 62 were included in this review after the screening process. A Preferred Reporting Items for Systematic Reviews and Meta-Analyses flow diagram reports the results of our search and filter strategy (figure 1).
A Preferred Reporting Items for Systematic Reviews and Meta-Analyses flow diagram illustrating the search and filter strategy.
The ‘normal’ gut microbiome
Several studies have examined the gut microbiome in ‘normal’ individuals with variations noted among populations.9 10 This is not surprising as age, geographic location, genetics and diet have all been shown to affect gut microbiome composition.11 12 Overall, the human gut microbiome is predominately constituted by the phyla Firmicutes, Bacteroides, Proteobacteria and Actinobacteria.9 13 14 These four phyla were dominant in the gut of 17 healthy individuals living in France or the Netherlands,10 as well as in 39 healthy individuals spanning six nationalities (Danish, Spanish, Italian, French, American and Japanese).9 At the genus level however, greater variation exists in the microbiome. For example, Faecalibacterium, Ruminococcus and Eubacterium were the most common genera in France and the Netherlands,10 while Bacteroides, Prevotella and Ruminococcus were the most abundant genera among the six aforementioned nationalities.9 Geographic variations in microbiome composition have also been noted in other regions. For example, in a study of 326 children (0–17 years) and 202 adults, significant phylogenetic differences were noted between both children and adults living in Malawi, Venezuela, and the USA, with US residents having the least diverse microbiome.12
Overall, it is difficult to tease out which underlying contributors (genetic and/or environmental) most robustly contribute to geographic variability in microbiome composition. In addition to differences in geography, weather and genetics, Malawians and Venezuelans have diets dominated by maize-derived, cassava-derived and other plant-derived polysaccharides, whereas US children and adults have diets richer in readily absorbed sugars.12 In fact, studies have shown that diet alone can influence microbiome composition. One study investigated the effect of a lacto-ovo-vegetarian diet for 3 months on the gut microbiome of 15 healthy individuals who were omnivorous previously. After 3 months of the lacto-ovo-vegetarian diet, the abundance of Alistipes was reduced, while an increased abundance of Roseburia inulinivorans, Ruminococcus lactis, Lactobacillus plantarum and Streptococcus thermophiles was noted.15 This study suggests that even short-term dietary changes may modify the gut microbiome.
Age and genetics also affect microbiome composition. In one study, the gut microbiome of 12 unrelated children and 1 fraternal twin pair was regularly sampled during their first year of life. From the first stool produced after birth through approximately 6 months, significant microbiome variability existed between the children. However, from approximately 6 months to 1 year, their microbiomes migrated toward a similar ‘adult-like’ composition with a predominance of Firmicutes and Bacteroidetes.11 Interestingly, the twin pair harboured a more similar microbiome compared with the other infants. More support for the impact of genetics and diet on microbiome composition comes from a Canadian study that examined 173 white and 182 South Asian 1-year-old infants whose mothers all lived in Canada for an average of 8 years. South Asian mothers were more likely to be vegetarian compared with white mothers. South Asian infants were found to have an increased abundance of Bifidobacterium, Atopobium, Streptococcus and Enterococcus and a decreased abundance of Firmicutes compared with white infants. This study suggests that the gut microbiome composition of infants may be influenced by genetics and both the infants’ and mothers’ diet.16 Collectively, these studies highlight that various factors may affect the gut microbiome and that there is variation with respect to what constitutes a ‘normal’ microbiome.
The gut microbiome influences immune function
The gut microbiome influences immune system development and function. For example, among Swiss-Webster mice that were 3–6 months of age, germ-free mice had lower numbers of immunologically competent cells in the lymphoid tissue of the mesenteric, submaxillary, axillary and popliteal lymph nodes compared with conventionally house mice.17 Furthermore, germ-free mice were more susceptible to infection by bacteria,18 including Listeria monocytogenes,19 Salmonella enterica,20 Klebsiella pneumoniae21 and fungus (Cryptococcus).22 Interestingly, mice were not only susceptible to systemic infection but also to infections at peripheral sites, such as an increased susceptibility to Pseudomonas aeruginosa keratitis.23 Fortunately, the noted immune abnormalities and their functional consequences were overcome when the gut microbiome was restored by faecal transplant from healthy mice, including normalised proportions of T-helper-17 (Th17) and T-regulatory (Treg) cells.24 This suggests that the microbiome is critical in maintaining normal levels of various immune mediators, such as plasma cells, Th17 and Treg, which are important for protection against infections at multiple sites.
Via its effect on Th17 (proinflammatory) and Treg (anti-inflammatory) cell regulation, the gut microbiome can also impact inflammation in multiple compartments.25 26 For example, some Clostridia species can stimulate Th17 expansion in intestinal and extraintestinal sites (proinflammatory), while other Clostridia species produce short chain fatty acids (SCFA) that support Treg proliferation (anti-inflammatory). Similarly, some Bacteroides species express molecules such as polysaccharide A (PSA) which mediates the conversion of CD4+ T cells into Treg cells and suppresses the Th17 inflammatory response, thus permitting the colonisation of Bacteroides fragilis.27 28 In fact, oral administration of PSA was found to both prevent and improve experimental colitis in mice,29 highlighting the important role of B. fragilis in the gut as a facilitator of Treg differentiation which induces mucosal tolerance.27 Overall, the gut microbiome is essential for the development of a healthy gut immune system that protects against infection with pathogenic micro-organisms while dampening excessive proinflammatory T-cell responses.
Gut dysbiosis and autoimmune disease
Alterations in the gut microbiome, also known as gut dysbiosis, have been linked to a variety of diseases. However, different autoimmune diseases have different microbiome signatures.
For example, in one study, 50 individuals with RA were found to have decreased gut microbial diversity, including a lower abundance of common commensals such as Bifidobacteria and Bacteroidetes compared with 51 individuals with fibromyalgia (FM). Of note, individuals with FM were chosen as controls because while FM is a non-inflammatory condition, individuals with RA and FM often receive overlapping medications, such as non-steroidal anti-inflammatory drugs and have similar age and sex distributions.8 Another study involving 44 individuals with new-onset untreated RA found an increased abundance of Prevotella copri compared with 26 individuals with chronic-treated RA and 28 healthy individuals.30 Interestingly, the abundance of P. copri did not significantly differ in treated RA compared with healthy individuals, which suggests a two-way connection between the microbiome and inflammatory states in which treating the underlying disease may alter the gut microbiome composition. Beyond its presence, P. copri has also been demonstrated to have immune relevance in RA. Notably, in 17 of the 40 individuals with RA, a specific protein of P. copri was shown to stimulate effector T-cell responses.31 Moreover, in the same study, 41 of the 127 individuals with RA demonstrated IgG or IgA antibody reactivity with P. copri, however this finding was rare in individuals with other types of arthritis.31 These data suggest that gut dysbiosis may contribute to manifestations of immune-mediated diseases.
Outside of RA, individuals with other autoimmune diseases, including ones associated with ocular inflammation (ie, uveitis) and/or immune-mediated dry eye, have been found to have different microbiome signatures, however, with inconsistencies noted across the literature. For example, in 49 individuals with spondylarthrosis, a significant increase in Ruminococcus gnavus abundance was demonstrated as compared with 17 individuals with RA and 18 healthy controls.32 However, in a study involving 103 individuals with ankylosing spondylitis compared with 104 healthy controls, a decreased abundance of Ruminococcus was demonstrated along with an increased abundance of the families Veillonellaceae and Lachnospiraceae.6 In 22 individuals with Behcet’s, a significant depletion in the genera Roseburia and Subdoligranulum, two Clostridium clusters, was found when compared with 16 healthy controls.7 Yet in another study involving nine individuals with Behcet’s, the abundance of Bacteroides uniformis was significantly higher than in 9 healthy controls, but the significant bacterial differences found in the first study were not demonstrated.33 Finally, in a study involving 35 individuals who underwent allogeneic haematopoietic stem cell transplantation, patients microbiomes were sampled at the time of acute (aGVHD) onset and compared with 35 post-transplant individuals without GVHD. Interestingly, specific microbiome signatures were associated with gastrointestinal aGVHD severity. Specifically, negative correlations were noted between aGVHD severity and abundance of the anaerobic bacteria Lachnospiraceae and Ruminococcaceae.34 Overall, many factors may explain the noted inconsistencies between studies including differences in the studied populations (eg, genetics, diet, geography, control comparison group) and methodologic variability in profiling microbiome composition. For example, the two Behcet’s studies were conducted in different geographic locations, that is, Italy7 and Japan,33 making comparisons between studies challenging due to the potential for confounding variables.
Overall, gut dysbiosis has been described in several autoimmune diseases include including RA, spondylarthrosis and Behcet’s, however with differences in microbiome composition between studies, even within the same disease.7 8 32 35
The gut microbiome in Sjögren’s syndrome
Sjögren’s is a chronic autoimmune disease characterised by oral and ocular dryness. It affects 0.5%–4% of the population, including more than 2 million Americans living with the disease.36 Gut dysbiosis has also been found in Sjögren’s, both in mouse models and humans.
In mice, the gut microbiome has been found to drive ocular manifestations of dry eye, including corneal staining and T-cell profiles. In one study, mice received desiccating stress to the ocular surface through the use of a fan with or without scopolamine, a medication that reduces tear production. Desiccating stress alone altered the gut microbiome, with an increase in Proteobacteria, as compared with non-stressed mice. When mice received oral antibiotics with a desiccating stress, a greater change to the gut microbiome was noted, with reductions in the phyla Bacteroidetes and Firmicutes and an increase in Proteobacteria beyond that which occurred with desiccating stress alone. In addition to these microbiome changes, a more severe dry eye phenotype was observed, with more severe corneal staining compared with controls.35 This demonstrates the role of the gut microbiome in dampening stress-induced ocular surface inflammation.
The gut microbiome also impacts disease phenotype in an immune-mediated dry eye animal model. Specifically, CD25 knockout (KO) mice develop dry eye along with other manifestations of Sjögren’s syndrome. Germ-free CD25KO mice were found to have a worse dry eye phenotype when compared with CD25KO conventionally housed mice, including increased lacrimal gland inflammation, glandular destruction and interferon-gamma (IFN-γ) producing T-cell infiltration in the lacrimal gland.37 Interestingly, improving the gut microbiome in germ-free CD25KO mice through faecal transfer improved dry eye phenotype, with improved corneal barrier function, increased goblet cell density and decreased lymphocytic infiltration, CD4+IFN-γ cell number and expression of IFN-γ.
Furthermore, the interface between the microbiome and the immune system has been examined in experimental models. In one experiment, immunodeficient mice received T cells from either (a) germ-free CD25KO mice, (b) germ-free CD25KO that received a faecal transplant from healthy conventionally housed mice or (c) conventionally housed mice. The latter two groups had less severe dry eye signs (less corneal barrier disruption, less T-cell infiltration into the lacrimal gland, greater goblet cell density and decreased frequency of pathogenic CD4+IFN-γ cells) compared with the former group.38 This suggests that the gut microbiome influences the pathogenicity of CD4+ T cells, with CD4+ T cells being more pathogenic in mice that lack a gut microbiome.
Overall, these studies demonstrate that gut microbiome composition can modulate dry eye disease severity induced from internal or external stressors in mice. The gut microbiome also impacts the pathogenicity of immune cells, which when transferred may independently transmit disease. However, it is understood that mice models do not fully recapitulate the dry eye disease noted in humans. As such, human studies examining these questions are vital.
Gut dysbiosis has also been identified in humans with Sjögren’s, however similar to other autoimmune diseases, inconsistencies have been found across literature. One study compared 10 individuals with Sjögren’s living in Texas to 10 healthy controls selected from the Human Microbiome Project who were also from Texas. Individuals with Sjögren’s had a greater relative abundance of Pseudobutyrivibrio, Escherichia/Shigella and Streptococcus and reduced relative abundances of Bacteroides, Parabacteroides, Faecalibacterium and Prevotella. Moreover, an inverse relationship was found between microbiome diversity and disease severity.35 Another study examined 10 individuals with Sjögren’s dry eye (SDE), 14 individuals with evaporative dry eye (dry eye symptoms and tear break up time (TBUT)<10 s) and 12 healthy volunteers in Seoul, South Korea. Overall, individuals with SDE demonstrated decreased genera of Blautia, Dorea and Agathobacter compared with controls and increased genera of Prevotella, Odoribacter and Alistipes compared with the evaporative dry eye group. Additionally, National Eye Institute (NEI) Score was positively related to Bacteroidetes (R2=0.12, p=0.04) and negatively related to Bifidobacterium (R2=−0.12, p=0.04). Tear secretion was positively related to Actinobacteria (R2=0.33, p<0.001) and Bifidobacterium (R2=0.26, p=0.001). TBUT was positively related to Actinobacteria (R2=0.53, p<0.001) and Bifidobacterium (R2=0.40, p<0.001) and negatively related to Bacteroidetes (R2=−0.15, p=0.02). Interestingly, after adjustment for confounders, multivariate linear regression analysis revealed that tear secretion was related to Prevotella (β=0.26, p=0.03) and TBUT was related to both Prevotella (β=0.25, p=0.04) and Actinobacteria (β=0.66, p=0.001).39
Our group has also investigated microbiome signatures in immune-mediated dry eye. In one study, gut microbiome signatures were compared between 13 individuals with SDE, 8 individuals with dry eye symptoms and features of Sjögren’s but who did not meet full criteria (non-Sjögren’s dry eye (NDE); ≥1 early Sjögren’s marker positivity, aqueous tear deficiency or a comorbid autoimmune disease) living in South Florida and 21 healthy individuals whose stool samples were provided by the stool bank OpenBiome. On a phylum level, all subjects with dry eye had a depletion of Firmicutes and an expansion of Proteobacteria, Actinobacteria and Bacteroidetes compared with controls. Interestingly, the phylogenetic diversity was increased when comparing individuals with SDE compared with controls, independent of age or comorbid autoimmune diseases. Furthermore, across the spectrum from controls to NDE to SDE, there was a decrease in genera Faecalibacterium and Veillonella, classes Ruminococcaceae and Lachnospiraceae and orders Clostridiales and Bacteroides. There was also an increase in genera Megasphaera, Parabacteroides and Prevotella. Finally, changes in certain classes of bacteria were associated with dry eye symptoms (dry eye questionnaire (DEQ) 5 and ocular surface disease index (OSDI) scores) and signs (ocular surface inflammation, corneal staining and tear production) when adjusting for age, gender, ethnicity and race.40
In a follow-up study, 10 individuals with immune-mediated dry eye living in South Florida, 5 of whom met full Sjögren’s criteria, were compared with a single healthy control sampled 12 times (OpenBiome). Similar to our first study, individuals with dry eye had a greater phylogenetic diversity and a decreased abundance of the genera Faecalibacterium and Ruminococcus. Unlike the prior study however, a decreased abundance of Prevotella was observed in cases compared with the control.41
Examining these four studies, both consistencies and discrepancies emerge between studies. Two studies found increased abundances of Blautia and Streptococcus and reduced abundances of Faecalibacterium and Prevotella in individuals with dry eye compared with controls.35 41 However, one study found a decreased abundance of Blautia and an increased abundance of Prevotella.39 The differences in microbiome compositions among studies may be due to several factors, including the aforementioned population-based differences, variations in processing and analytical techniques for microbiome analysis or comorbid diseases. Further studies are needed that characterise the microbiome in Sjögren’s and examine potential confounders.
The impact of the gut microbiome on inflammatory cells and markers on immune-mediated dry eye and Sjögren’s
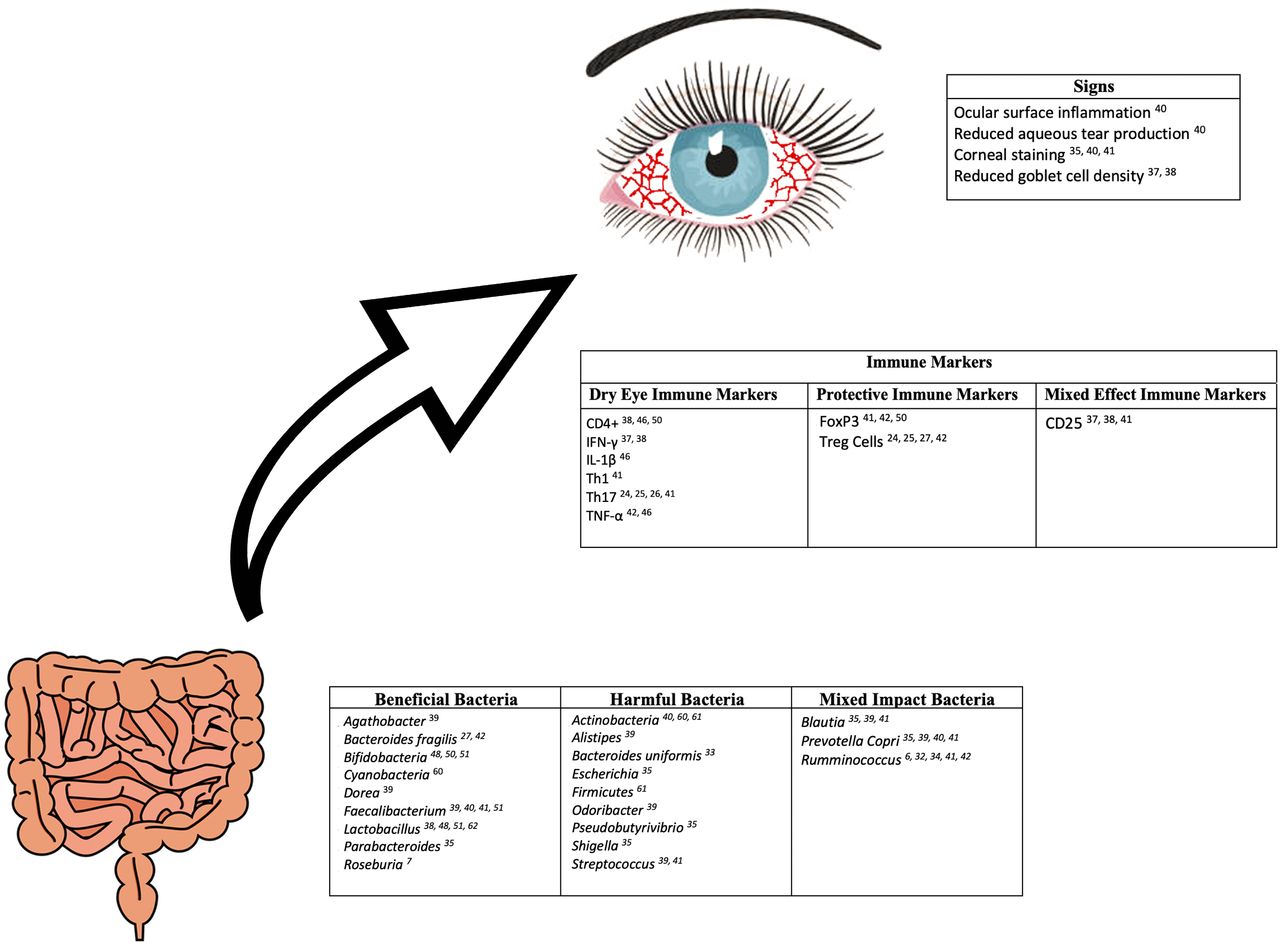
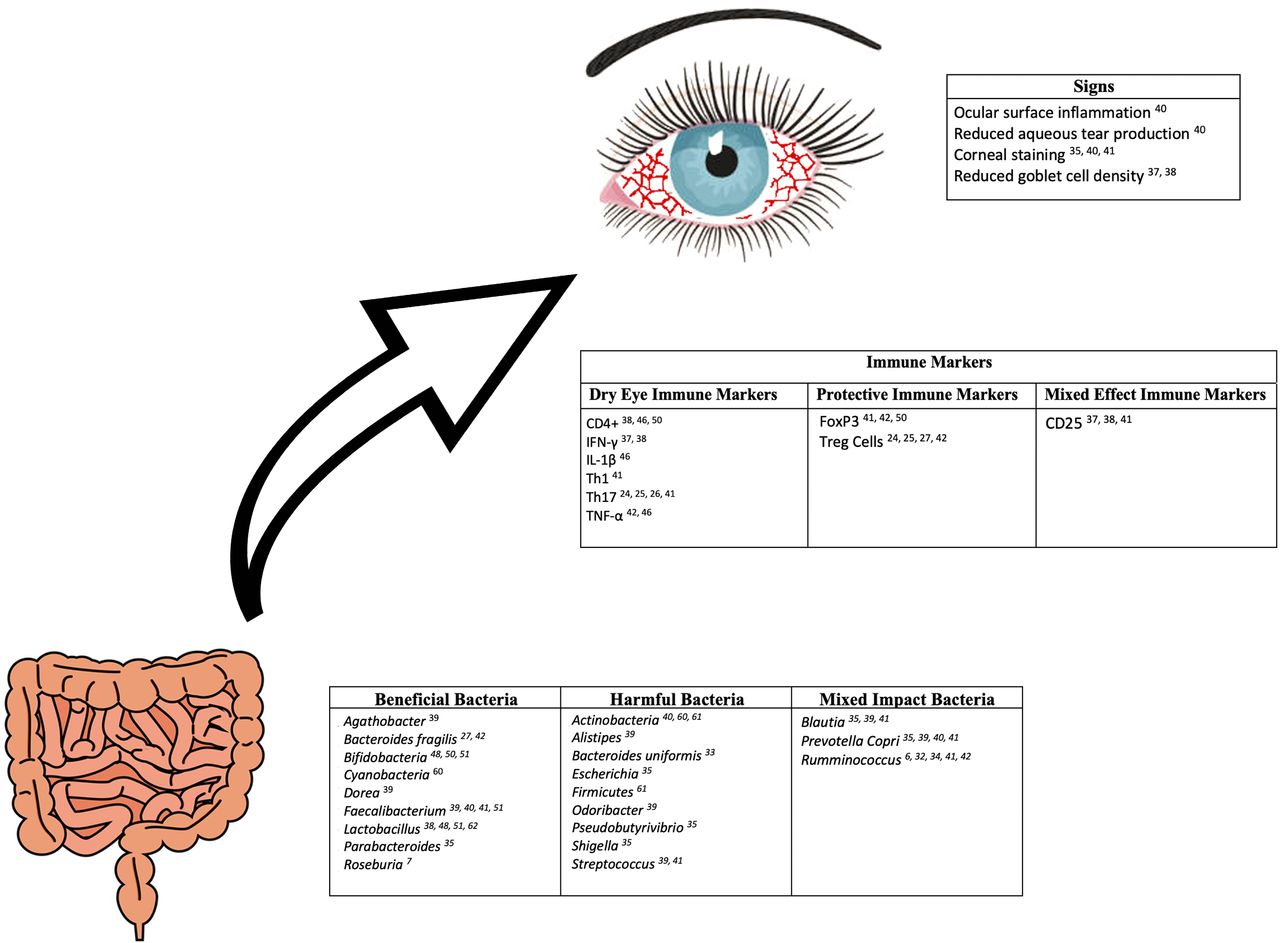
The association between the gut microbiome and T-cell profiles in immune-mediated dry eye and Sjögren’s has been explored in a few studies (figure 2). As mentioned previously, germ-free CD25KO mice were found to have a worse dry eye phenotype when compared with CD25KO conventionally housed mice.37 This suggests that gut dysbiosis contributes to increased lymphocyte-associated inflammation. Furthermore, in our study of individuals with immune-mediated dry eye, we demonstrated relationships between various dry eye metrics and T-cell profiles in blood. Dry eye symptom severity was positively related to both effector T cells (Th1: r=0.76, p=0.01; Th17: r=0.83, p=0.003) and regulatory T cells (CD25: r=0.66, p=0.04; FoxP3: r=0.68, p=0.03) frequencies.41 Positive associations were noted between corneal staining and effector T cells (Th1: r=0.48, p=0.19; Th17: r=0.47, p=0.21), while negative associations were identified between corneal staining and regulatory T cells (CD25: r=−0.66, p=0.06; FoxP3: r=−0.54, p=0.13). Although these latter relationships were not statistically significant potentially due to an underpowered analysis of only 10 individuals, these findings may suggest the severity of corneal staining is positively associated with effector T-cell populations and inversely associated with regulatory T-cell populations.41
{kind=link}
{kind=link}
Reported relationships between gut bacteria, immune markers, and signs of dry eye disease.
Relationships between soluble inflammatory markers and microbiome composition have also been examined in a Spanish study that included 19 individuals with Sjögren’s and 19 age-matched, sex-matched and body mass index-matched healthy controls.42 In patients with Sjögren’s, the proinflammatory cytokines interleukin-17 (IL-17) and tumor necrosis factor (TNF)-α were negatively associated with serum levels of Bifidobacterium (R2=0.742, B=−1.289, p<0.001 and R2=0.697, B=−0.580, p<0.001, respectively), while IL-17 was positively associated with Ruminococcus (R2=0.742, B=−0.815, p<0.001). IL-12 was negatively associated with Lachnospira (R2=0.897, B=−0.601, p<0.001), Roseburia (R2=0.897, B=−0.641, p<0.001) and Bifidobacterium (R2=0.897, B=−0.715, p<0.001) and positively associated with Enterobacter (R2=0.897, B=0.562, p<0.001). IL-6 was negatively associated with Blautia (R2=0.835, B=−0.132, p=0.022) and Roseburia (R2=0.835, B = −0.785, p=0.001) and positively with Escherichia coli (R2=0.835, B=0.678, p<0.001). This same pattern was not noted in healthy controls. Specifically, a significant negative association was only noted between the proinflammatory cytokines IL-6 and TNF-α and Parabacteroides distasonis (R2=0.948, B=1.139, p<0.001 and R2=0.570, B=0.570, p=0.011, respectively). Interestingly, both individuals with Sjögren’s and healthy controls shared the same anti-inflammatory cytokine associations. In both groups, the anti-inflammatory cytokine IL-10 was positively associated with Faecalibacterium prausnitzii (R2=0.325, B=0.570, p=0.011; R2=0.494, B=1.337, p<0.001, respectively) and Ruminococcus (R2=0.325, B=0.259, p=0.029; R2=0.494, B=0.259, p=0.029, respectively). Finally, FOXP3 mRNA expression, which is implicated in the development and function of Treg cells, was positively associated with B. fragilis in both individuals with Sjögren’s (R2=0.547, B=0.548, p=0.029) and healthy controls (R2=0.547, B=0.670, p=0.016).42 Collectively, these studies suggest that gut dysbiosis is associated with alterations in cellular and soluble immune profiles in immune-mediated dry eye. While larger studies across diverse populations are necessary to validate the noted correlations between bacterial composition, clinical metrics and immune profiles, the presence of such associations suggest that modulating the microbiome may serve as a therapy for immune-mediated dry eye.
Therapeutically targeting the microbiome
Gut microbiome manipulation, with the goal of restoring a healthy microbiome, has been examined as an approach to alter disease phenotype in autoimmune diseases. Strategies that have been explored in this regard include diet modification, probiotics, prebiotics and faecal microbial transplant (FMT) (table 1).
Gut microbiome manipulation via diet
The role of diet and nutrition has been investigated as a therapy for immune-mediated dry eye. One study examined relationships between a Mediterranean diet and Sjögren’s. In 82 adults diagnosed with primary Sjögren’s and 51 adults who had symptoms but did not meet the criteria for Sjögren’s, higher Mediterranean diet scores (calculated via a questionnaire based on the amount of certain food intake per week) were associated with a lower odds of having Sjögren’s. Interestingly, the strongest inverse associations were noted with fish and vegetable consumption on multivariate analysis.43 Fish intake has also been shown to reduce the risk of other autoimmune diseases such as RA.44 Although no data specifically exist on how diet affects the microbiome in individuals with immune-mediated dry eye, it is increasingly recognised that diet may have an impact on health by impacting gut microbiome composition.45
Animal studies have also examined the role of diet in dry eye. In one study, the impact of a high-fat diet on lacrimal gland function was investigated in 4-week-old C57BL/6 mice. The results showed that mice on a high-fat diet demonstrated pathologic changes, including lower levels of tear secretion, increased inflammatory CD4+ T cell infiltration, increased proinflammatory factors such as TNF-α and IL-1β and increased apoptosis of acinar and myoepithelial cells compared with mice with standard diets. After a standard diet was introduced to the mice previously on a high-fat diet, pathologic changes in the lacrimal gland were partially reversed, including a reduction in inflammatory cells and proinflammatory factors and an upregulation of anti-inflammatory cytokines.46 These studies suggest a relationship between certain diets, disease severity and inflammatory signatures.
Gut microbiome manipulation via probiotics and prebiotics
Probiotics and prebiotics have also been studied as a way to improve gut microbiome composition and downstream metabolites in diseases associated with immune-mediated dry eye. One prospective clinical trial evaluated the efficacy of prebiotics in preventing symptoms and signs of acute GVHD. Prebiotic supplements that contained resistant starch, glutamine, fibre and oligosaccharides were administered daily to 43 individuals before their allogenic haematopoietic stem cell transplantation until 28 days after transplantation, while 142 individuals did not receive prebiotics before their transplantation. Overall, prebiotics reduced the incidence of acute GVHD of all grades 100 days after transplant (53% vs 73%; p=0.004). In addition, individuals who received prebiotics had a reduced duration (median, 11 days vs 14 days; p<0.001) and severity (measured by Eilers’ Oral Assessment Guide) of oral mucositis, a shortened duration of diarrhoea (median, 7 days vs 9 days; p=0.049) and a shortened duration of opioid use (median 8 days vs 10 days, p=0.013) compared with controls. Moreover, a greater frequency of individuals in the prebiotic versus control group had stable or increased microbial alpha diversity (Shannon index) at day 28 compared with baseline (increased: 20% vs 2.8%; stable, 23.3% vs 20.8%, decreased, 57.7% vs 76.4%; p=0.004).47
Probiotics have also been studied with respect to RA, a disease often comorbid with secondary Sjögren’s. In a double-masked study of 60 individuals with RA, half received probiotic capsules containing Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium bifidum, whereas the other half received a cellulose-filled placebo capsule. After 8 weeks of daily intervention, probiotic supplementation significantly improved RA disease activity scores and decreased the inflammatory marker high-sensitivity C-reactivity protein concentrations.48
In addition, the effects of prebiotics and probiotics on dry eye have been evaluated in animal models. One study found that mice with acute GVHD had significantly reduced levels of the SCFA butyrate in their intestinal epithelial cells compared with controls.49 Replenishing butyrate levels either through direct administration of butyrate as a prebiotic or by supplying butyrate-producing Clostridia strains as a probiotic significantly improved intestinal epithelial cell junctional integrity, decreased their apoptosis and reduced acute GVHD severity in mice. Unfortunately, dry eye parameters were not specifically reported in this study.49
The disease modifying effects of prebiotics and probiotics may be due to their anti-inflammatory properties. In one randomised double-blinded study in Ireland, 27 healthy volunteers who received prebiotics containing Bifidobacterium infantis were compared with 12 healthy volunteers who received a placebo. The prebiotics group demonstrated an increased IL-10, an anti-inflammatory cytokine and increased numbers of Foxp3+CD4+T cells, important players in mucosal immune tolerance, in peripheral blood.50 This study suggests that B. infantis leads to immunoregulatory responses, highlighting its potential benefit individuals with inflammatory disease. These findings may translate to individuals with immune-mediated dry eye, however studies on the effects of prebiotics and probiotics on individuals with immune-mediated or SDE have yet to be reported.
Gut microbiome manipulation via FMT
FMT is the process of transferring intestinal microbiome from a healthy donor to a patient with the goal of replacing an abnormal gut microbiome with a stable and healthy one. Despite the technical difficulties, FMT has been explored as a therapy for autoimmune diseases associated with immune-mediated dry eye. For example, in an open-label study, one or two doses of FMT (from healthy spouses or relatives) were delivered by enema to four individuals with GVHD after stem cell transplantation. One month after FMT, improved gastrointestinal symptoms including defecation consistency and frequency were noted. This correlated with an increase in beneficial bacteria including Lactobacillus, Bacteroides, Bifidobacterium and Faecalibacterium.51 However, no data were given on FMT’s effect on dry eye in this publication.
In a study by our group, the safety and efficacy of FMT in 10 individuals with immune-mediated dry eye, 5 of whom met full Sjögren’s criteria and 5 of whom had early markers of Sjögren’s, were investigated. After recipients received two FMTs via enema, 1 week apart, from one healthy donor (OpenBiome), eight of the ten microbiomes migrated towards the donor microbiome. By the 3-month follow-up however, most individuals regressed back to their native microbiome. Notably, certain phyla, classes and genera abundances remained more similar to the donor profile as compared with the recipients’ baseline profiles out to 3 months. Ultimately, no adverse effects were identified with the use of FMT, and half of the study cohort (four of the five individuals who met Sjögren’s criteria) subjectively reported improved gastrointestinal and dry eye symptoms at last follow-up. Despite this, overall, dry eye symptoms and signs were statistically unchanged with therapy.41
FMT has also been investigated in other autoimmune diseases, including ulcerative colitis, multiple sclerosis and idiopathic thrombocytopenic purpura. In a randomised control trial, 75 individuals with ulcerative colitis received six weekly FMTs from healthy donors or placebo. The remission frequency at 7 weeks (24% vs 5%) was significantly higher in the FMT versus placebo groups.52 FMT has also been examined in individuals with metabolic syndrome, in a study that highlighted the importance of donor characteristics in influencing FMT’s effect. In a double-masked study of 18 obese individuals (body mass index>30 kg/m2 or waist circumference>102 cm with fasting plasma glucose level>5.6 mmol/L), subjects were randomised to allogenic (from lean male donors) or autologous (from own collected faeces) FMT via a gastroduodenal tube. Increased sensitivity to insulin was noted in recipients 6 weeks after a single dose of FMT from lean donors but not after autologous transplant.53 Furthermore, FMT has been explored as a treatment for autoimmune neurological conditions. In an open-label study of three individuals with multiple sclerosis and constipation, daily FMT infusions (route not reported) for 1–2 weeks resulted in an objective improvement in neurological symptoms, including regaining the ability to walk, decreased paresthesia and improved energy levels.54 With FMT being successful in other autoimmune conditions, gut microbiome manipulation via FMT remains a potential future therapy for immune-mediated dry eye, with the most effective route and frequency still yet to be determined.
The role of the OSM in immune-mediated dry eye
The OSM may also have a role in immune-mediated dry eye. Of note, the OSM is paucibacterial compared with the gut, with 0.06 bacteria per human conjunctival cell compared with the gut which has 10 bacterial cells for every one human cell.55 56 The composition of the OSM also differs from the gut microbiome, with Proteobacteria dominating on the ocular surface as opposed to Firmicutes in the gut.57 58
Studies have examined the inferior conjunctival OSM in dry eye in varying populations using 16S rDNA sequencing, with discrepant results. One study examined the OSM in 15 individuals with SDE versus eight healthy controls living in Texas. Individuals with Sjögren’s were not observed to have any significant differences with regards to microbiome composition, richness or structure.35 However, findings across the literature have not been uniform, with several studies reporting OSM diversity and compositional differences in individuals with dry eye. One study conducted in Beijing, China, examined 23 individuals with SDE, 36 patients with NDE and 39 healthy controls. Unlike the Texas study, individuals with Sjögren’s had a significantly decreased alpha diversity compared with controls and NDE, as well as significant compositional differences (beta diversity) between the three groups. Most pronounced differences included a significantly increased relative abundance of Actinobacteria in the SDE group compared with both the NDE group and controls and a significantly decreased Cyanobacteria and Bacteroides abundance in the SDE group compared with controls.59 Similarly, in a South Korean study that examined the OSM in 48 individuals with primary Sjögren’s compared with 72 subjects with NDE, a significantly reduced alpha diversity was noted in the Sjögren’s group compared with NDE. On the other hand, significant compositional differences were not identified between the groups.60 Overall, while reductions in alpha diversity were noted in the Sjögren’s versus other groups in the two Asian studies, beta diversity findings were not uniform.
Studies have also reported OSM differences in immune-mediated dry eye not limited to Sjögren’s. One study in Shaanxi, China, divided individuals with dry eye (OSDI≥13 and Schirmer’s≤5) into groups by the presence (n=38) or absence (n=49) of an autoimmune disease (including systemic lupus erythematosus, RA, Sjögren’s, systemic sclerosis, Graves). Unlike the Asian studies, individuals with autoimmune disease had a similar alpha diversity but significant compositional differences compared with those without an autoimmune disease, most notably increased relative abundances of Actinobacteria, Firmicutes and Bacteroides and a significantly decreased relative abundance of Proteobacteria.61
Taken together, differences in OSM composition have been described in individuals with immune-mediated dry eye compared with non-immune dry eye subtypes and controls, however with discrepancies between studies regarding alpha diversity and compositional differences. As with gut microbiome composition, potential confounders such as population-based differences (geography, diet, demographics, comorbid diseases) may explain the divergent findings across studies. While larger studies across diverse populations needed, the OSM may also be a therapeutic target for immune-mediated dry eye through microbial modulation.
OSM manipulation in immune-mediated dry eye
Modulating the OSM may be a potential therapy in dry eye; however, research exploring this concept is sparse. One study evaluated the efficacy of Lactobacillus eye drops on symptoms (itching, photophobia, burning, tearing) and signs (conjunctival hyperemia, chemosis, secretion, Trantas dots, superficial punctuate keratitis) in individuals with vernal keratoconjunctivitis. In an open label study in Italy, seven patients were treated Lactobacillus eye drops four times per day for 4 weeks in both eyes. After 4 weeks of treatment, a significant improvement in symptoms and signs was reported in six patients, most notably in symptoms of photophobia, itching, tearing and signs of conjunctival hyperemia and chemosis.62 While these results are promising, it is unknown if these findings will translate into beneficial effects in immune-mediated dry eye. As such, further studies are needed.
Conclusion
Overall, the studies above highlight complex interactions between microbiomes, the immune system and immune-mediated dry eye. A balance between proinflammatory and anti-inflammatory bacterial species is important for immune system development and function, to protect the body locally and systemically from infection and to appropriately dampen inflammatory response. Several studies have found gut microbiome alterations in autoimmune diseases associated with immune-mediated dry eye, and as a result, the gut microbiome has emerged as a potential therapeutic target to mitigate symptoms and signs of immune-mediated dry eye via dietary manipulation, prebiotics and probiotics and FMT. However, as there is not one apparent microbial signature in immune-mediated dry eye, it is not clear which therapy is optimal in which individual. Furthermore, the role of the OSM in immune-mediated dry eye is less well understood and as such, is an important avenue for future studies.
Ethics statements
Patient consent for publication
Ethics approval
Not applicable.
References
Footnotes
Contributors All authors have contributed to the idea, literature review, writing, editing and review of the manuscript.
Funding This study was supported by the Sjögren's Foundation, the University of Miami Interdisciplinary Team Science Award (UM SIP 2018-2R), the Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development, Clinical Sciences R&D I01 CX002015 (AG) and Biomedical Laboratory R&D Service I01 BX004893 (AG), Department of Defense Gulf War Illness Research Program W81XWH-20-1-0579 (AG) and Vision Research Program W81XWH-20-1-0820 (AG), National Eye Institute R01EY026174 (AG) and R61EY032468 (AG), NIH Center Core Grant P30EY014801 (institutional) and Research to Prevent Blindness Unrestricted Grant (institutional).
Competing interests None declared.
Provenance and peer review Commissioned; externally peer reviewed.