Article Text

Abstract
Background We aim to determine the possible adverse effects of ciprofloxacin (CPFX) and tetracycline (TETRA), as examples of bactericidal and bacteriostatic agents, respectively, on cultured human retinal pigment epithelial cells (ARPE-19).
Methods Cells were treated with 30, 60 and 120 µg/mL of CPFX and TETRA. Cell metabolism was measured by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. JC-1 dye (5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide) assay was conducted to measure the mitochondrial membrane potential (MMP). The level of reactive oxygen species (ROS) was measured using the -2’,7’-dichlorodihydrofluorescein diacetate assay (H2DCFDA). Quantitative real-time PCR was performed to analyse the gene expression levels associated with apoptosis (BAX, BCL2-L13, BCL2, Caspase 3, Caspase 7 and Caspase 9), inflammatory (interleukin-1β (IL-1β), IL-6, IL-33, transforming growth factor-α (TGF-α), TGF-β1 and TGF-β2) and antioxidant pathways (SOD2, SOD3, GPX3 and NOX4), along with the mitochondrial DNA (mtDNA) copy numbers.
Results Results illustrated that while all three concentrations of CPFX decreased cellular viability of ARPE-19 during all incubation periods, the 120 µg/mL TETRA resulted in increased cellular viability. At 48 and 72 hours, levels of MMP and ROS decreased significantly with each antibiotic. BAX, BCL2-L13, CASP-7, CASP-9, SOD2 and GPX3 genes overexpressed by either antibiotics. There was higher expression of IL-6 and IL-1B with TETRA treatment. The level of mtDNA decreased using both treatments.
Conclusions Clinically relevant concentrations of CPFX and TETRA have detrimental impacts on ARPE-19 cell lines in vitro, including upregulation of genes related to apoptosis, inflammation and antioxidant pathways. Additional studies are warranted to investigate if these harmful effects might be seen in retinal degeneration models in vivo.
- retina
- apotosis
- drugs
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
This subject is relatively new, this study examines the possible adverse effects of ciprofloxacin (CPFX) and tetracycline (TETRA).
Bactericidal and bacteriostatic antibiotics might induce detrimental impacts on retinal cell lines, notably in the population suffering from age-related disease.
What are the new findings?
Our new findings include clinically adjusted dosages of CPFX and TETRA may have detrimental impacts human retinal pigment epithelial (ARPE-19) cells.
How might these results change the focus of research or clinical practice?
We speculate that if CPFX can have deleterious effects on the wild type, healthy ARPE-19 cells, then it may also have deleterious effects on damaged mitochondria of patients with age-related disease such as age-related macular degeneration who are exposed to treatments with the fluoroquinolones.
Introduction
Fluoroquinolones (ie, ciprofloxacin (CPFX), ofloxacin, levofloxacin, fleroxacin, lomefloxacin, gatifloxacin) are currently a popular group of bactericidal antibiotics used to treat (1) skin, urinary tract, joint, sinus and lung infections; (2) ocular infections, such as endophthalmitis and bacterial keratitis1 2 and (3) traumatic injuries prophylactically. Fluoroquinolones preliminarily inhibit the DNA gyrase (topoisomerase II) enzyme, which is involved in supercoiling, separation and replication of circular bacterial DNA.3 Fluoroquinolones are very effective against intracellular, gram negative and positive organisms. Moreover, the fluoroquinolone penetrance through ocular barriers increases its impact for eye diseases.4
Bacteriostatic tetracyclines (TETRA) are efficient in treating skin, urinary, respiratory and chlamydia/trachoma infections. These antibiotics impede bacterial protein synthesis, which eventually prevent further bacterial growth and replication. TETRA bind to the bacterial 30S ribosome, preventing aminoacyl tRNA from interacting with the ribosome RNA complex. Furthermore, these antibiotics may alter the bacterial cytoplasmic membrane, causing leakage of cell material, thereby facilitating cell death.5 While antibiotics have significant preventative and therapeutic effectiveness clinically, studies suggest that some antibiotics have significant adverse effects. For instance, with respect to the eye, there are increased risks of retinal detachment, optic neuritis and retinal haemorrhage associated with administrations of fluoroquinolones.6 Other adverse effects of antibiotics include permanent damage to the inner ear cells (auditory and vestibular), tendon damage and rupture, arthropathy, destruction of kidney cells and psychosis.7 Human mammary epithelial cells (HMEC) in vitro show elevated levels of reactive oxygen species (ROS), protein carbonylation, lipid peroxidation and 8-hydroxy-2’2deoxyguanosine (marker for DNA damage) after treatment with fluoroquinolones.8 These features were also present in mice treated 16 weeks with the antibiotics.8 Primary human osteoblasts (PHO), osteosarcoma and HeLa cells were impaired by fluoroquinolone treatments.9 High-content screening for mitochondrial proteins showed that several fluoroquinolones damaged the mitochondria of human liver cells.10
In 2015, over 32 million prescriptions for fluoroquinolones were given to patients in the USA for various medical conditions. Anticancer studies report that CPFX can halt cell cycle and cause double-strand DNA breaks that lead to increased apoptosis.11 Exposure to CPFX causes lower cell viability and induces apoptosis in lung, melanoma and hepatocellular cancer cell lines.12 CPFX blocks topoisomerase II inhibition in malignant cells but not normal cells13 and induces G2 cell cycle arrest,14 ultimately suggesting that a course of CPFX may be a reasonable adjunct therapy for some cancers.15 While most people do not have any serious side effects, a small percentage have progressive, severe complications. Case reports of serious damage to multiple systems, including peripheral neuropathies, muscle weakness, pain in joints and tendons, cognitive impairment, along with gastrointestinal and respiratory disturbances, were shown for individuals treated with levofloxacin.16 More recently, an excellent review on the negative impact of fluoroquinolones for a small number of individuals was published.17 In 2015, the Food and Drug Administration (FDA) recognised a syndrome called fluoroquinolone-associated disability (FQAD) to describe otherwise healthy subjects that took fluoroquinolones and subsequently developed irreversible, severe side effects.
Mitochondria originated from ancestral aerobic bacteria,18 19 and present-day bacteria and mitochondria possess many structural and biological similarities, such as similar outer membrane proteins and genomic sequence.20 21 Therefore, it is not surprising that both would be detrimentally impacted by antibiotics. In particular, it is critical to determine if these antibiotics have negative influence on mitochondria from elderly patients that already have compromised mitochondrial functions. For example, studies have shown that patients with age-related macular degeneration (AMD) possess damaged and dysfunctional mitochondria that have increased susceptibility to stressors.22 23 In AMD retinas, the retinal pigment epithelial (RPE) cells are the first cell type affected in this disease. In this study, we investigated how CPFX and TETRA affect mitochondrial and cellular health in human ARPE-19 cells.
Materials and methods
Cell culture
ARPE-19 cells were purchased from the American Type Culture Collection (Manassas, Virginia, USA) and grown in a mixture of Dulbecco’s Modified Eagle’s medium/nutrient mixture F-12 (Invitrogen, Carlsbad, California, USA), 10% fetal bovine serum, 0.37% sodium bicarbonate, 0.58% L-glutamine, antibiotics (streptomycin sulphate 0.1 mg/mL, amphotericin-B 2.5 mg/mL, penicillin G 100 U/mL and gentamycin 10 mg/mL) and 10 mM non-essential amino acids. Cells were incubated in standard conditions (95% humidity, 5% CO2 at 37°C).
Cells were treated with either CPFX (Cat#17850, Sigma-Aldrich, St. Louis, Missouri, USA) or TETRA (Cat# 87128, Sigma-Aldrich) at a of 0, 30, 60 and 120 µg/mL and cultured for 24, 48 and 72 hours. We used a hydrochloric acid (HCl) solution with 0.1 normality and methanol (Meth) as the vehicles for CPFX and TETRA, respectively.
Cell metabolism (MTT assay)
Cell metabolism levels were measured with the MTT assay. ARPE-19 cells were cultured in 96-well plates (104/well) and 10 µL MTT assay reagent (3-(4,5-dimethyltiazol-2-yl)−2,5-dipheniltetrazolium bromide) (Catalogue# 30006, Biotium, California, USA) was added to each well and plates were incubated at 37°C for 2 hours. Then, 100 µL/well DMSO was added to each well and plates were read in an absorbance reader (signal at 570 nm and reference at 630 nm) (Biotek Elx808 Absorbance Reader, Winooski, Vermont, USA). Experiments were performed three times. There were 12 replicate wells for each treatment modality.
Reactive oxygen species (ROS assay)
Cells were cultured at a density of 104/well in 96-well plates. Subsequently, 2’, 7’-dichlorodihydrofluorescein diacetate; Catalogue# D399, Thermo Fisher Scientific, Waltham, MA solution, which is converted into a fluorescent molecule in the presence of ROS, was added to each well and plates were read on a fluorescent plate reader (SoftMax Pro, V.6.4, Catalogue# 94089, Sunnyvale, California, USA) at excitation (EX, 492 nm) and emission (EM, 520 nm) wavelengths. There were 12 wells for each treatment modality. Experiments were performed three times. There were 12 replicate wells for each treatment modality.
Mitochondria membrane potential (ΔΨm)
Cells were seeded in 96-well plates (104/well). After the treatment periods, the JC-1 reagent (5,5’,6,6’- tetrachloro1,1’,3,3’-tetraethyl-benzimidaz olylcarbocyanine iodide; Catalogue# 30001, Biotium, California, USA) was added to each well and plates were incubated at 37°C for 15 min. Finally, plates were read on a fluorescent plate reader (SoftMax Pro, V.6.4, Catalogue# 94089,) at red (EX 550 nm and EM 600 nm) and green (EX 485 nm and EM 535 nm) emissions to determine the ratios of red to green fluorescence. Experiments were performed three times. There were 12 replicate wells for each treatment modality.
RNA/DNA isolation and cDNA amplification
ARPE-19 cells were treated with 120 µg/mL treatment concentrations of CPFX and TETRA. Subsequently, DNA and RNA were isolated from the cellular lysate via application of Pure Genomic DNA Mini Kit (Thermo Fisher Scientific; Cat#K1820-01,) and RNeasy Mini-Extraction kit (Qiagen), according to manufacturer’s protocol. The Nano Drop 1000 (Thermo-scientific) was used to determine the RNA/DNA concentration and purity from 15 samples of the ARPE-19 cells. Each RNA sample (100 ng) was reverse transcribed into cDNA using the QuantiTect reverse transcription kit (Qiagen).
Quantitative real time polymerase chain reaction (qRT-PCR)
We evaluated the expression levels of genes related to the apoptosis (BAX, BCL2-L13, BCL2, Caspase 3, Caspase 7 and Caspase 9 (mitochondria specific)) (QuantiTect Primer Assay, Qiagen), inflammatory markers (interleukin-1β (IL-1β), IL-6, IL-33, transforming growth factor-α (TGF-α), TGF-β1 and TGF-β2) and antioxidant enzymes (SOD2, SOD3, GPX3 and NOX4) (table 1) by quantitative real-time-PCR (qRT-PCR). The total RNA was isolated from cultured treated cells and vehicle-control cells. Then QuantiFast SYBR Green PCR Kit (Qiagen, USA) on a Bio-Rad iCycler detection system was used for the Q-PCR. The HPRT1 primer was used as the housekeeping gene and standardisation of expression levels for all primers. Analyses were done in triplicates and no template control wells were used to assess the contamination. For all the experiments, HPRT1 gene was selected as the housekeeping gene. ΔΔCt method used for analysing the obtained data of qRT-PCR, which ΔCt = [Ct (threshold value) of the target gene] − [Ct for HPRT1], and ΔΔCt = ΔCt of the treatment condition − ΔCt of the untreated condition. The fold changes of treated conditions compared with untreated condition were calculated as: fold change=2−ΔΔCt.
Information of the genes related to apoptotic, inflammatory and antioxidant pathways
Mitochondrial DNA copy number assay
The relative levels of mitochondrial DNA (mtDNA) copy numbers were measured for all samples by comparing the levels of mtDNA (MT-ND2) versus nuclear DNA (18S). The total DNA was isolated from the cultured treated cells, untreated and vehicle-control cells. TaqMan Gene Expression assay (Thermo Fisher Scientific, USA) was performed for the quantitative measurement of mtDNA. Analyses were done in triplicates.
Statistical analyses
Using GraphPad Prism (V.5.0, GraphPad Software., and San Diego, California, USA) was used for all statistical analyses. The data was analysed by two-way analysis of variance with the BONFERRONI test. P<0.05 was considered statistically significant.
Results
Cell viability
Performed MTT assay revealed the mean percentages of cellular metabolism in cultures treated for 24 hours with 60 and 120 µg/mL concentrations of CPFX decreased by 21.33% (p=0.04) and 28.4% (p=0.014), respectively, compared with the vehicle-control cells. After 48 hours of incubation, viability was reduced by 20.79% (p=0.025) with 30 µg/mL, 27.85% (p=0.018) with 60 µg/mL and 34.57% (p=0.011) with 120 µg/mL CPFX in comparison with vehicle-control samples. Moreover, in the 72 hours treatment period, the mean percentages declined by 27.46% (p=0.03) with 30 µg/mL, 40.68% (p=0.014) with 60 µg/mL and 45.55% (p=0.01) with 120 µg/mL compared with vehicle-control samples (figure 1A).
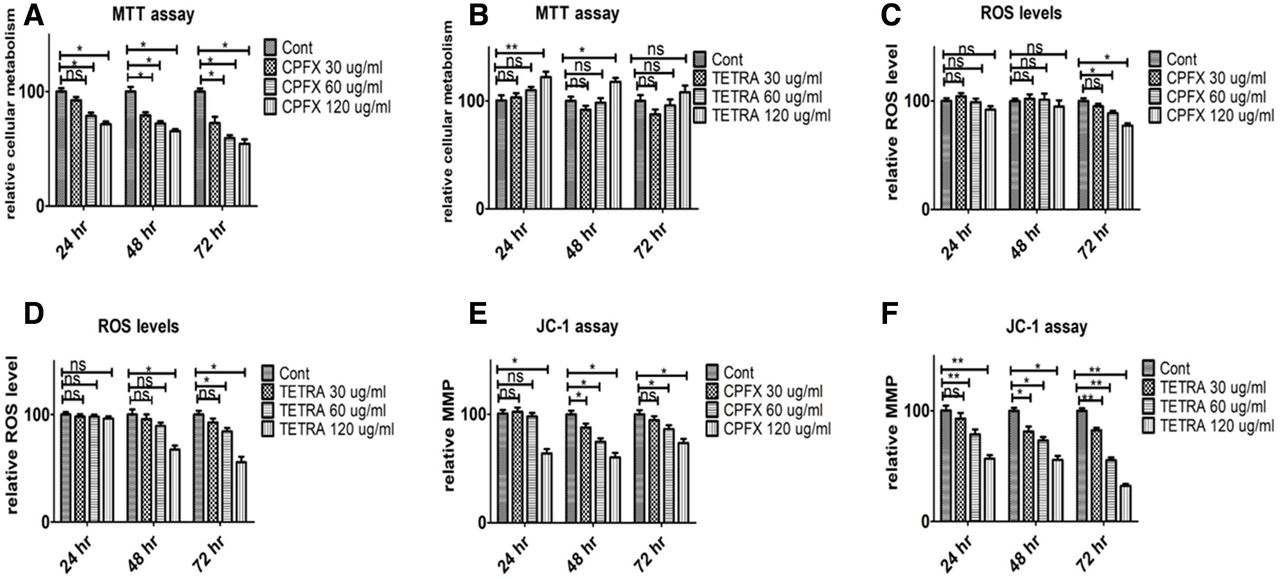
Treatment effects of CPFX and TETRA on cellular metabolism (figure 1A,B), ROS levels (figure 1C,D) and mitochondrial membrane potential (MMP, figure 1E,F) in ARPE-19 cells after 24, 48 and 72 hours as measured with the MTT, H2DCFDA and JC-1 assays. *P<0.05, **P<0.01. ARPE-19, human retinal pigment epithelial cells; CPFX, ciprofloxacin; H2DCFDA, 2’,7’-dichlorodihydrofluorescein diacetate; ns, not significant; ROS, reactive oxygen species; TETRA, tetracycline.
In 24 hours cultures with TETRA, the mean percentage of cell viability of ARPE-19 cells treated with 120 µg/mL TETRA increased by 22.1% (p=0.001) compared with vehicle-control wells (figure 1B). After a 48-hour treatment period at 120 µg/mL TETRA, there was a 17.7% (p=0.049) increase in the mean percentage of cellular metabolism compared with the vehicle-control cells.
Altogether, these findings demonstrated that there is a negative association between the higher concentrations of CPFX and cellular viability of ARPE-19 cells, notably in prolonged exposure periods. However, high concentrations of TETRA may induce positive influential effects on the viability of ARPE-19 cells.
ROS production
The ROS levels in CPFX-treated cultures at 24 and 48 hours showed no significant changes compared with vehicle-control samples (figure 1C). However, after 72 hours culture with either CPFX 60 or 120 µg/mL, the mean percentages of ROS levels declined by 11.27% (p=0.048) and 22.59% (p=0.028), respectively, in comparison with vehicle-control cells.
TETRA-treated ARPE-19 cultures showed similar results on ROS levels (figure 1D). After 48 hours, cells treated with TETRA 120 µg/mL had a 32.6% (p=0.031) decrease in ROS levels compared with untreated samples. Notably, at 72 hours, the mean percentage of ROS levels decreased by 15.62% (p=0.023) with TETRA 60 µg/mL and 44.36% (p=0.032) with TETRA 120 µg/mL compared with vehicle-control cells. These results suggest that higher concentrations of both antibiotics decrease ROS levels, most notably after longer incubation times.
Changes in mitochondrial membrane potential
Compared with vehicle-control cells, the mean percentage of mitochondrial membrane potential (MMP) diminished by 36.81% (p=0.031) in samples treated 24 hours with 120 µg/mL CPFX. The MMP was reduced by 12.08% (p=0.037) with CPFX 30 µg/mL, by 25.27% (P=0.032) with CPFX 60 µg/mL and by 39.68% (p=0.013) with CPFX 120 µg/mL treatment after 48 hours culture compared with untreated samples. The results showed that cells treated with 60 and 120 µg/mL CPFX for 72 hours had 11.27% (p=0.035) and 22.59% (p=0.025) decrease of mean percentages in comparison with vehicle-control cells (figure 1E). The MMP mean percentages of cells treated with 60 and 120 µg/mL TETRA for 24 hours were reduced by 21.61% (p=0.021) and 43.58% (p=0.014), respectively, compared with vehicle-control cultures. Furthermore, cells cultured for 48 hours showed a reduction by 18.6% with 30 µg/mL (p=0.026), 26.75% with 60 µg/mL (p=0.021) and 44.26% with 120 µg/mL (p=0.014) compared with vehicle-control cultures. There was also a decrease in ΔΨm after 72 hours of TETRA treatment; with mean percentages decline by 17.44% (p=0.003) with 30 µg/mL, 44.51% (p=0.004) with 60 µg/mL and 67.69% (p=0.001) with 120 µg/mL, respectively, compared with vehicle-control wells (figure 1F). Altogether, these in vitro findings show that higher concentrations of both CFX and TETRA decreased MMP over time.
Expression levels for pro-apoptosis, antioxidant and pro-inflammation genes
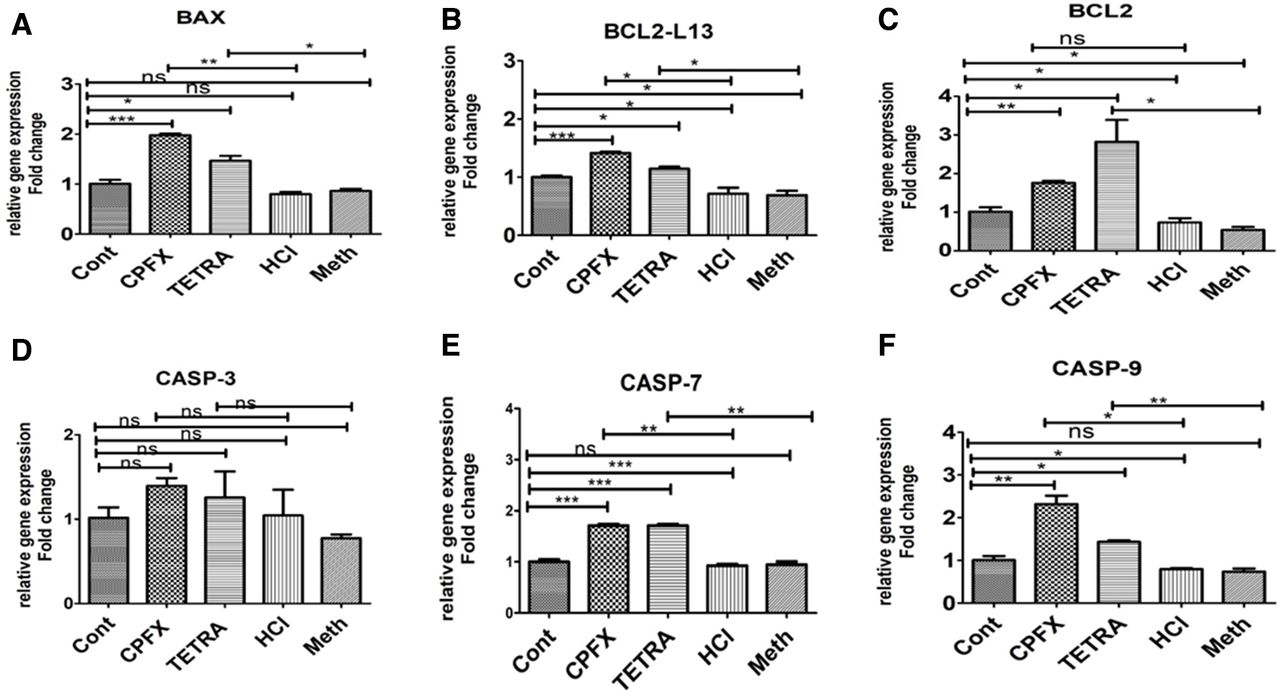
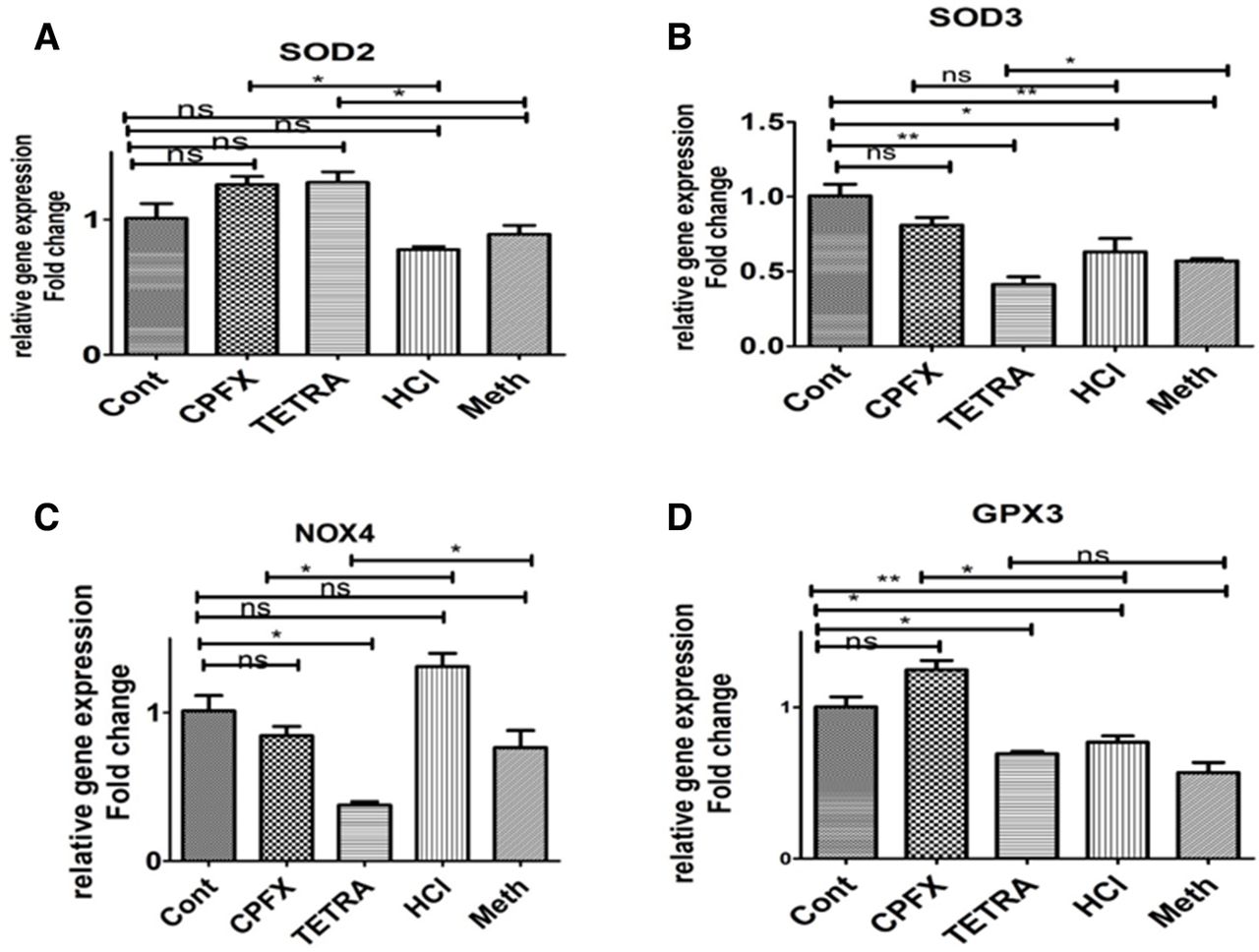
In all experiments of the qRT-PCR, the vehicle-control cells were considered at a value of 1 (table 2). After treatment with CPFX, the relative gene expression of BAX (1.977-fold, p=0.002) (figure 2A), BCL2-L13 (1.414-fold, p=0.014) (figure 2B), Caspase 7 (1.712-fold, p=0.003) (figure 2E), Caspase 9 (2.319-fold, p=0.013) (figure 2F), SOD2 (1.26-fold, p=0.01(figure 3A) and GPX3 (1.24-fold, p=0.018) (figure 3D) were increased as compared with the HCl-treated control cells. However, the relative gene expression levels of NOX4 in CPFX exposed cells decreased to 0.84-fold (p=0.031) compared with HCl-treated controls (figure 3C).
Expression levels in ARPE-19 Treated cells with CPFX and TETRA compared with vehicle-treated controls

Expression of apoptotic genes (A) BAX, (B) BCL2-L13, (C) BCL-2, (D) CASP-3, (E) CASP-7 and (F) CASP-9 in CPFX and TETRA treated ARPE19 cells. *P<0.05, **P<0.01, ***P<0.001. ARPE19, human retinal pigment epithelial cells; Cont, untreated control; CPFX, ciprofloxacin; HCl, hydrochloric acid vehicle control; Meth, methanol vehicle control; ns, not significant; TETRA, tetracycline.
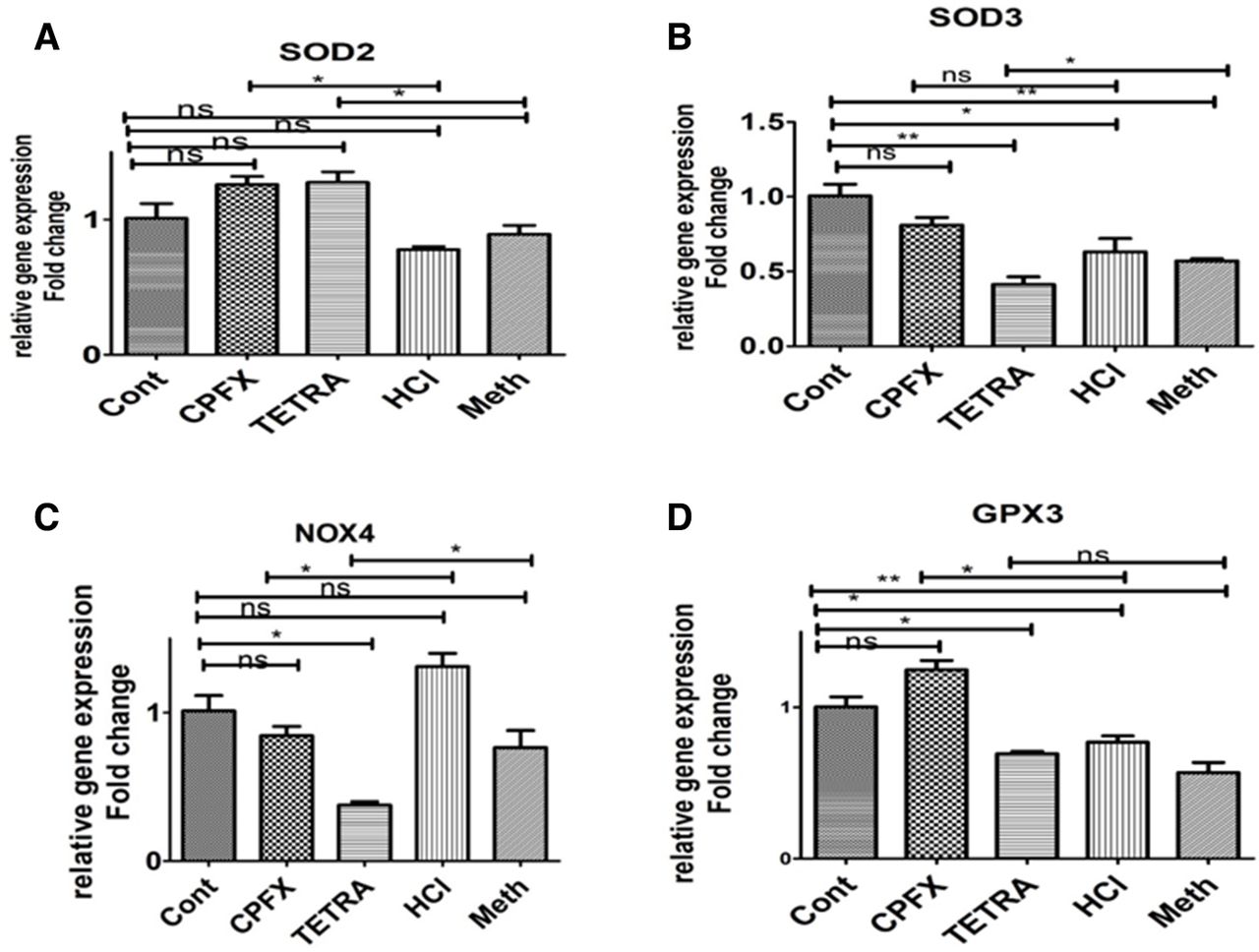
Expression of antioxidant genes (A) SOD2, (B) SOD3, (C) NOX4 (D) GPX3 in CPFX and TETRA treated ARPE19 cells. *P<0.05, **P<0.01, ***P<0.001. ARPE19, human retinal pigment epithelial cells; Cont, untreated control; CPFX, ciprofloxacin; HCL, hydrochloric acid vehicle control; Meth, methanol vehicle control; ns, not significant; TETRA, tetracycline.
The TETRA-treated cells had relative gene expression levels of BAX (1.465-fold, p=0.04) (figure 2A), BCL2-L13 (1.142-fold, p=0.026) (figure 2B), BCL2 (2.82-fold, p=0.024) (figure 2C), Caspase-7 (1.712-fold, p=0.002) (figure 2E), Caspase-9 (1.434-fold, p=0.007) (figure 2F) and SOD2 (1.27-fold, p=0.048) (figure 3A) compared with the Meth-treated control group. Moreover, the relative gene expression levels of SOD3 and NOX4 in 120 µg/mL TETRA-treated cells decreased to 0.41-fold (p=0.041) (figure 3B) and 0.37-fold, (p=0.045) (figure 3C) in comparison with the Meth-treated control cells.
Although the relative gene expression of inflammatory marker IL-6 in CPFX-treated cells increased to 1.31-fold (p=0.048) (figure 4A), the relative gene expression of IL-33 declined to 0.34-fold (p=0.047) (figure 4C) compared with HCl-treated control cells. Moreover, in TETRA-treated cells, the relative gene expression of TGF-α1 (0.41-fold, p=0.014) (figure 4D), TGFβ1 (0.54-fold, p=0.043) (figure 4E) and IL-33 (0.11-fold, p=0.011) (figure 4C) compared with Meth-treated control samples. Also, in 120 µg/mL TETRA-treated cells, the relative gene expression of IL-6 and IL-1β increased to 5.1-fold (p=0.001) (figure 4A) and 2.29-fold (p=0.023) (figure 4B), compared with the Meth-treated control cells.

Expression of inflammatory pathway genes (A) IL-6, (B) IL-1β, (C) IL-33, (D) TGF-α1, (E) TGF-β1 and (F) TGF-β2 in CPFX and TETRA treated ARPE19 cells. *P<0.05, **P<0.01, ***P<0.001. ARPE19, human retinal pigment epithelial cells; Cont, untreated control; CPFX, ciprofloxacin; HCl, hydrochloric acid vehicle control; IL-6, interleukin-6; Meth, methanol vehicle control; ns, not significant; TETRA, tetracycline; TGF-β1, transforming growth factor-β1.
Therefore, it is possible that exposure of ARPE-19 cells with CPFX and TETRA facilitated higher expression of some apoptotic, inflammatory and antioxidant enzyme-related genes.
Significant reduction of mtDNA copy number
In CPFX-treated ARPE-19 cultures, the level of mtDNA decreased to 0.23-fold (p=0.008) compared with HCl-treated control cells (figure 5). In addition, treatment with TETRA reduced the relative level of mtDNA to 0.28-fold (p=0.041) in comparison with Meth-treated control cells (figure 5). The untreated cultures were assigned a value of 1. Our findings show that treatment of ARPE-19 with CPFX and TETRA led to significant reduction of mtDNA copy numbers.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of TETRA and CPFX on the of mtDNA copy number level compared with nDNA. *P<0.05, **P<0.01. Cont, untreated control; CPFX, ciprofloxacin; HCL, hydrochloric acid vehicle control; Meth, methanol vehicle control; mtDNA, mitochondrial DNA; ns, not significant; TETRA, tetracycline.
Discussion
In this study, we evaluated the effects of CPFX and TETRA on ARPE-19 cells in vitro. We assessed the outcomes of these treatments on levels of cellular viability, ROS, MMP and expression levels of genes involved in inflammatory, apoptotic and antioxidant enzymes pathways. Our findings showed that treatment with clinically relevant dosages of CPFX resulted in significantly decreased ARPE-19 cell viability, metabolic activity and MMP that were more prominent with longer incubation and higher concentrations of CPFX. Four of the proapoptosis genes (BAX, BCL2-L13, Casp-7 and Casp-9) were upregulated after CPFX treatment, which is consistent with the lower cell numbers. Additionally, IL-6 levels were higher in the CPFX-treated cells, indicating that elevated levels of inflammation would be found associated with CPFX treatment. None of the other inflammation-related genes (TGF-α, TGF-β1, TGF- β2 or IL1α) changed expression levels on CPFX exposure.
In the ARPE-19 cultures, ROS levels remained unchanged at the 24 and 48 hours time points with the 60 and 120 µg/mL CPFX dosages by 72 hours. These data suggest that the ARPE-19 response to CPFX was via the mitochondria (lower MMP) and the cellular metabolic activity through the Nicotinamide adenine dinucleotide phosphate (NADPH) flux (lower MTT) and not through the generation of ROS. The lower ROS levels observed in the CPFX and TETRA treated ARPE-19 cells may also in part be due to the upregulation of SOD2, and the downregulation of NOX4, which is responsible for the production of ROS. Moreover, other reports have shown enhanced lactate production due to stimulated glycolysis,24 which would also lead to lower electron transport chain (ETC) activity and decreased ROS production with reduced mitochondrial activity.
In contrast to our findings, Kalghatgi et al evaluated CPFX-treated non-cancer mammary epithelial cells (MCF10A), primary human aortic endothelial cells (PAEC), primary HMEC, human gut epithelial cells and normal human diploid skin fibroblasts and reported higher levels of ROS, lipid peroxidation and oxidative damage to DNA and proteins. Usually bactericidal antibiotics disrupt the ETC and tricarboxylic cycle which would lead to lower formation of the endogenously generated ROS.8
Many of the published data related to CPFX have been conducted using various cancer cell lines so it might be expected that our findings with ARPE-19 cells would be different. Cancer-related studies have illustrated CPFX has inhibitory effects on leukaemia cells, glioblastoma cells, colorectal and bladder cancer cells that act via diminished MMP and increased apoptosis.25–28 Duewelhenke et al evaluated the effects of 20 antibiotics on PHO, the osteosarcoma cell lines (MG63) and Hela epithelial cell lines demonstrating fluoroquinolones, including CPFX, decreased the metabolic activity and proliferation of PHO by 20% and inhibited mitochondrial oxidative phosphorylation, but the cells underwent a metabolic shift to use glycolysis, as measured by lactate concentrations.9 Similarly, Holtom et al analysed murine osteoblast-like cell lines (MC3T3-E1) and reported a 50% inhibitory concentration of 40 µg/mL CPFX at both 48 and 72 hours and decreased 5-bromo-2'-deoxyuridine at 80 µg/mL CPFX.29 In this study, we illustrated that similar harmful effects are found in normal, non-cancer RPE cells.
Fluoroquinolones, including CPFX, can induce considerable harmful influences on a variety of cancer cells. For instance, bladder cancer cells exposed to 100 µg/mL CPFX showed an upregulation of BAX, the proapoptotic regulator, and arrested S/G2-M phase of the cell cycle.27 Kloskowski et al reported similar results when they exposed three different cancer cell lines (lung cancer, melanoma and glioblastoma) to a range of concentrations (12–1092 µg/mL) of CPFX for 24–48 hours and they found that CPFX induced cell cycle arrest at the G2/M phase.13 This was supported in CPFX-treated melanoma cells (COLO829) that showed inhibited DNA polymerase II and arrest of the S checkpoint of cell cycle.30
In the ARPE-19 cells, the long-term incubation with higher dosages of TETRA resulted in increased cell viability and metabolism along with lower ROS levels, suggesting positive effects on cellular health with decreased MMP. Our data suggest that the overall effect of TETRA on the ARPE-19 cells is less disruptive than CPFX. Similarly, a study by Kalghatgi et al reported significant CPFX-induced damaged mitochondrial morphology, along with lower levels of MMP, ATP production and metabolic activity in treated cells.8 However, the same dosage of TETRA did not induce significant alterations of these mitochondrial-related features or affect ROS production31 compared with untreated group.
Our gene expression studies showed that ARPE-19 cells had a differential response to TETRA with upregulation of the antiapoptosis gene BCL2, downregulation of TGF-β1 (growth and differentiation) and higher levels of IL-1β, a proinflammatory gene compared with the CPFX-treated cells. However, in a Zebrafish model (Danio rario), Ding et al found that TETRA induced damage at several concentrations (45, 60 and 90 mg/L) after 7, 14 and 21 days with mitochondrial damage, including diminished mitochondrial cristae and mitochondrial swelling.32 Another study in hepatocytes showed significant negative influences on mitochondrial calcium uniporter with treatment after TETRA derived compounds (50 uM, 22.22 g). TETRA prevented Ca2+ uptake by mitochondria and then inhibited the inducement of mitochondrial permeability transmission by Ca2+.33
Conclusions
Future investigations including in vivo studies and clinical trials are necessary in the discovery of effective treatments with antibiotics with nominal adverse effects. In conclusion, clinically adjusted dosages of CPFX and TETRA may have detrimental impacts ARPE-19 cells. We speculate that if CPFX can have these deleterious effects on the wildtype, healthy ARPE-19 cells, then it may also have deleterious effects on the older, damaged mitochondria of the AMD subjects are exposed to treatments with the fluoroquinolones.
Acknowledgments
This work was supported by the Discovery Eye Foundation, Polly and Michael Smith, Edith and Roy Carver, Iris and B. Gerald Cantor Foundation, Max Factor Family Foundation, and NEI R01 EY0127363 (MCK). Supported in part by an Unrestricted Departmental Grant from Research to Prevent Blindness. We acknowledge the support of the Institute for Clinical and Translational Science (ICTS) at University of California Irvine.
References
Footnotes
Contributors NS planned the study, designed experiments, wrote the manuscript, and analysed data. LS and AN edited the manuscript. MCK, the PI, developed the concepts, edited the manuscript, provided resources for the study. KS, SA, MC and AB helped with the experimental designs and performed some of the experiments. NS is an Arnold and Mabel Beckman Retinal Degeneration Fellow. KS is a Genentech AMD Research Fellow.
Funding This work was supported by the Discovery Eye Foundation, Polly and Michael Smith, Edith and Roy Carver, Iris and B. Gerald Cantor Foundation, Max Factor Family Foundation, and NEI R01 EY0127363 (MCK). Supported in part by an Unrestricted Departmental Grant from Research to Prevent Blindness. We acknowledge the support of the Institute for Clinical and Translational Science (ICTS) at University of California Irvine.
Competing interests None declared.
Patient and public involvement Patients and/or the public were not involved in the design, or conduct, or reporting, or dissemination plans of this research.
Patient consent for publication Not required.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article.